

ACTB Loss-of-Function Mutations Result in a Pleiotropic Developmental Disorder
- PMID: 29220674
- PMCID: PMC5812896
- DOI: 10.1016/j.ajhg.2017.11.006
ACTB Loss-of-Function Mutations Result in a Pleiotropic Developmental Disorder
Abstract
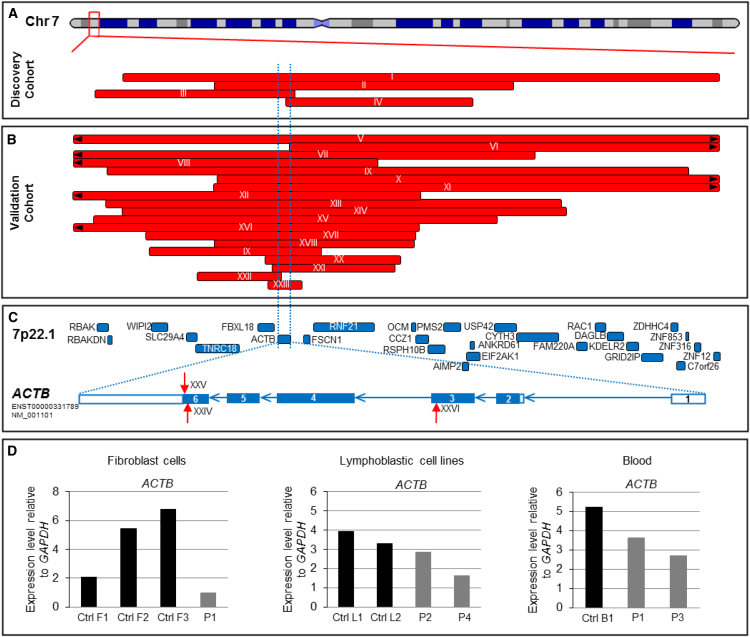
ACTB encodes β-actin, an abundant cytoskeletal housekeeping protein. In humans, postulated gain-of-function missense mutations cause Baraitser-Winter syndrome (BRWS), characterized by intellectual disability, cortical malformations, coloboma, sensorineural deafness, and typical facial features. To date, the consequences of loss-of-function ACTB mutations have not been proven conclusively. We describe heterozygous ACTB deletions and nonsense and frameshift mutations in 33 individuals with developmental delay, apparent intellectual disability, increased frequency of internal organ malformations (including those of the heart and the renal tract), growth retardation, and a recognizable facial gestalt (interrupted wavy eyebrows, dense eyelashes, wide nose, wide mouth, and a prominent chin) that is distinct from characteristics of individuals with BRWS. Strikingly, this spectrum overlaps with that of several chromatin-remodeling developmental disorders. In wild-type mouse embryos, β-actin expression was prominent in the kidney, heart, and brain. ACTB mRNA expression levels in lymphoblastic lines and fibroblasts derived from affected individuals were decreased in comparison to those in control cells. Fibroblasts derived from an affected individual and ACTB siRNA knockdown in wild-type fibroblasts showed altered cell shape and migration, consistent with known roles of cytoplasmic β-actin. We also demonstrate that ACTB haploinsufficiency leads to reduced cell proliferation, altered expression of cell-cycle genes, and decreased amounts of nuclear, but not cytoplasmic, β-actin. In conclusion, we show that heterozygous loss-of-function ACTB mutations cause a distinct pleiotropic malformation syndrome with intellectual disability. Our biological studies suggest that a critically reduced amount of this protein alters cell shape, migration, proliferation, and gene expression to the detriment of brain, heart, and kidney development.
Keywords: ACTB; chromatin; developmental disorder; malformations; β-actin.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures



References
-
- Yagi H., Furutani Y., Hamada H., Sasaki T., Asakawa S., Minoshima S., Ichida F., Joo K., Kimura M., Imamura S. Role of TBX1 in human del22q11.2 syndrome. Lancet. 2003;362:1366–1373. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
