

Regulation of Crossover Frequency and Distribution during Meiotic Recombination
- PMID: 29222342
- PMCID: PMC6542265
- DOI: 10.1101/sqb.2017.82.034132
Regulation of Crossover Frequency and Distribution during Meiotic Recombination
Abstract
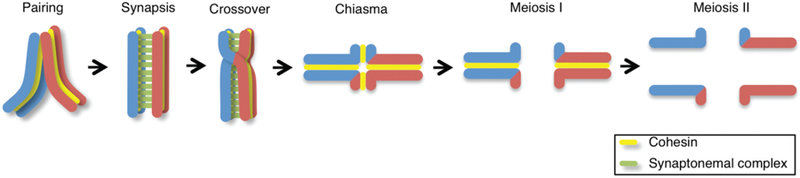
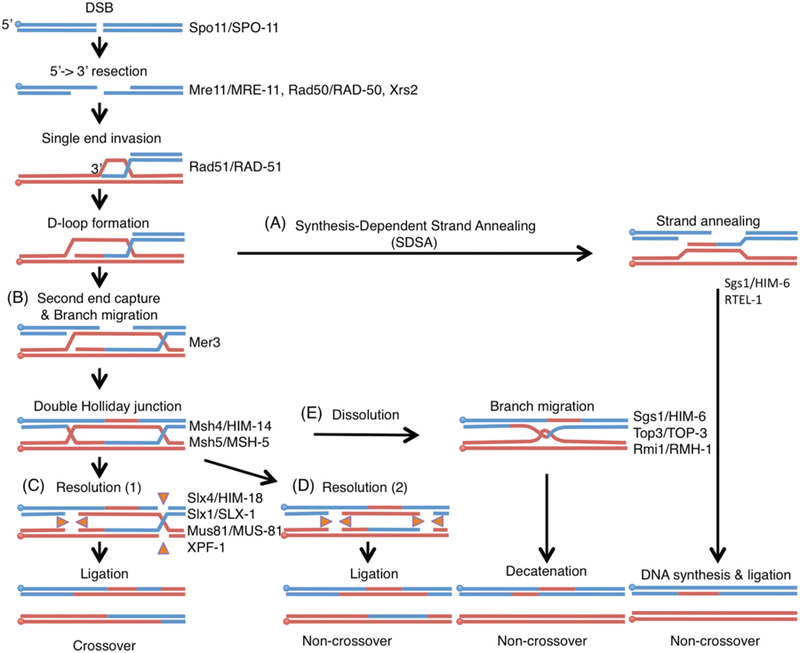
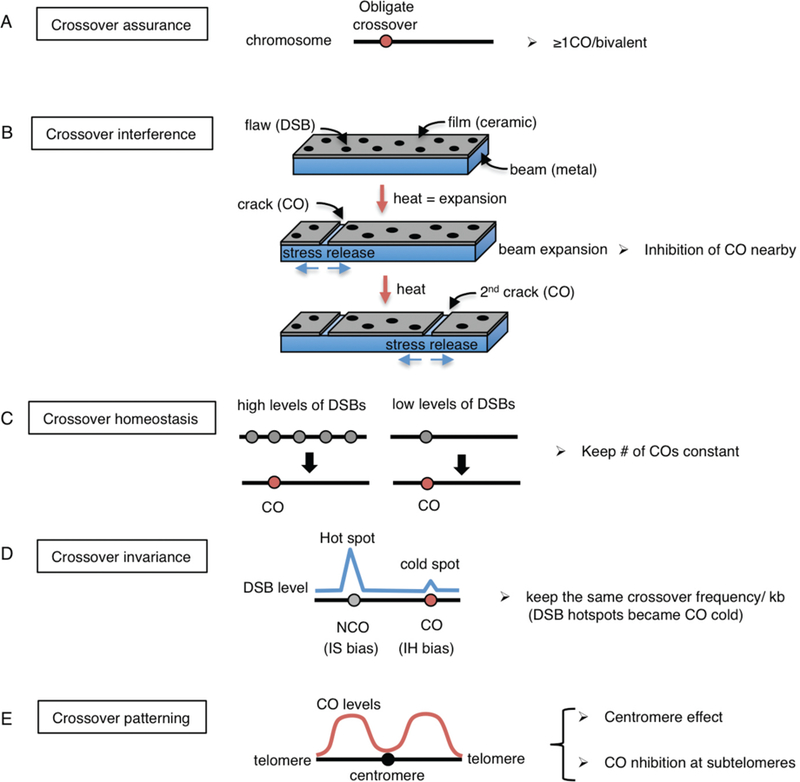
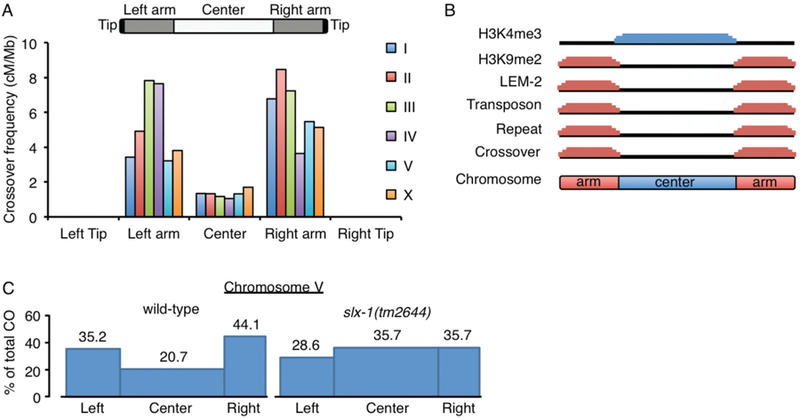
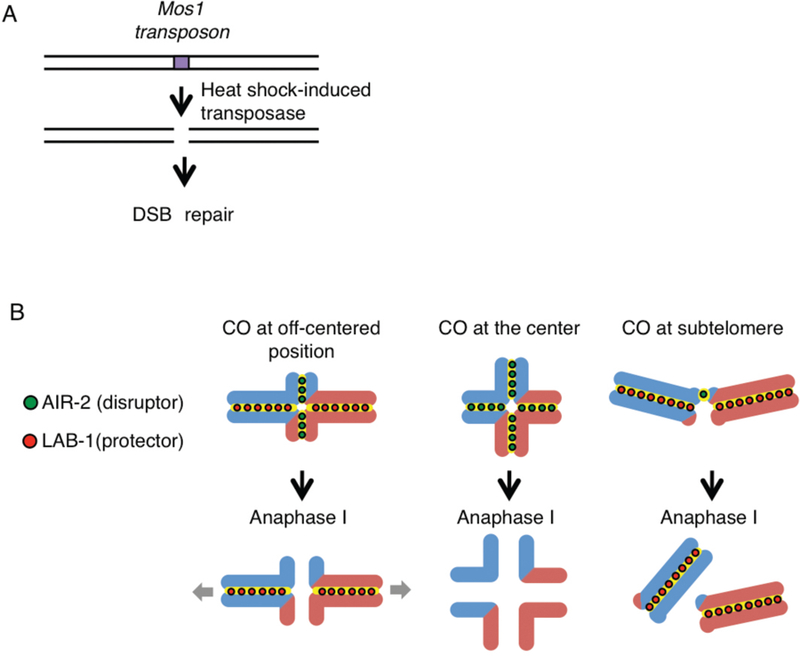
Crossover recombination is essential for generating genetic diversity and promoting accurate chromosome segregation during meiosis. The process of crossover recombination is tightly regulated and is initiated by the formation of programmed meiotic DNA double-strand breaks (DSBs). The number of DSBs is around 10-fold higher than the number of crossovers in most species, because only a limited number of DSBs are repaired as crossovers during meiosis. Moreover, crossovers are not randomly distributed. Most crossovers are located on chromosomal arm regions and both centromeres and telomeres are usually devoid of crossovers. Either loss or mislocalization of crossovers frequently results in chromosome nondisjunction and subsequent aneuploidy, leading to infertility, miscarriages, and birth defects such as Down syndrome. Here, we will review aspects of crossover regulation observed in most species and then focus on crossover regulation in the nematode Caenorhabditis elegans in which both the frequency and distribution of crossovers are tightly controlled. In this system, only a single crossover is formed, usually at an off-centered position, between each pair of homologous chromosomes. We have identified C. elegans mutants with deregulated crossover distribution, and we are analyzing crossover control by using an inducible single DSB system with which a single crossover can be produced at specific genomic positions. These combined studies are revealing novel insights into how crossover position is linked to accurate chromosome segregation.
© 2017 Saito and Colaiácovo; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Acquaviva L, Szekvolgyi L, Dichtl B, Dichtl BS, de La Roche Saint Andre C, Nicolas A, Geli V 2013. The COMPASS subunit Spp1 links histone methylation to initiation of meiotic recombination. Science 339: 215–218. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials