

Different carotenoid conformations have distinct functions in light-harvesting regulation in plants
- PMID: 29222488
- PMCID: PMC5722816
- DOI: 10.1038/s41467-017-02239-z
Different carotenoid conformations have distinct functions in light-harvesting regulation in plants
Abstract
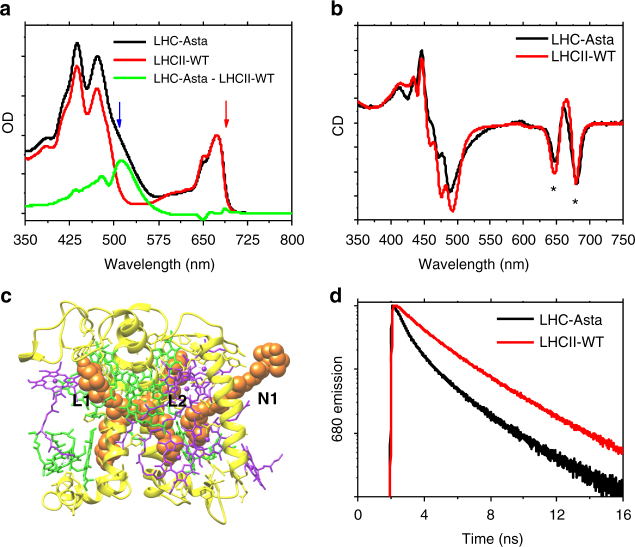
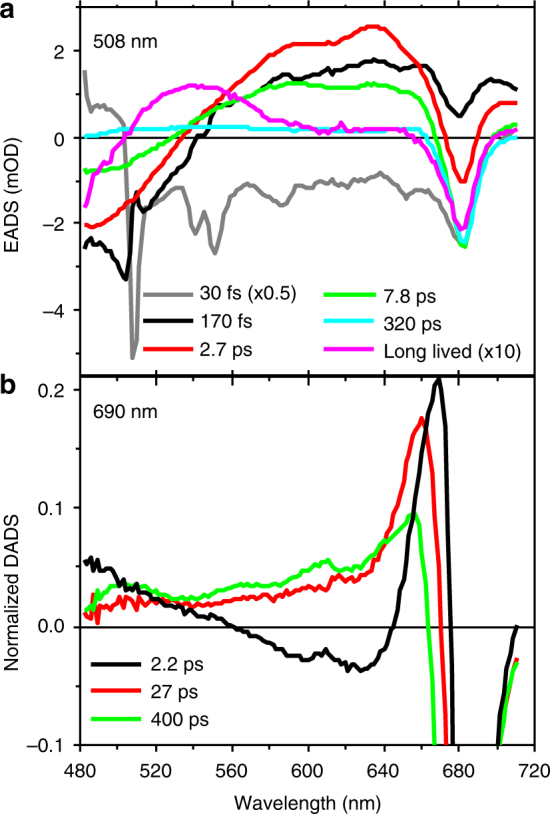
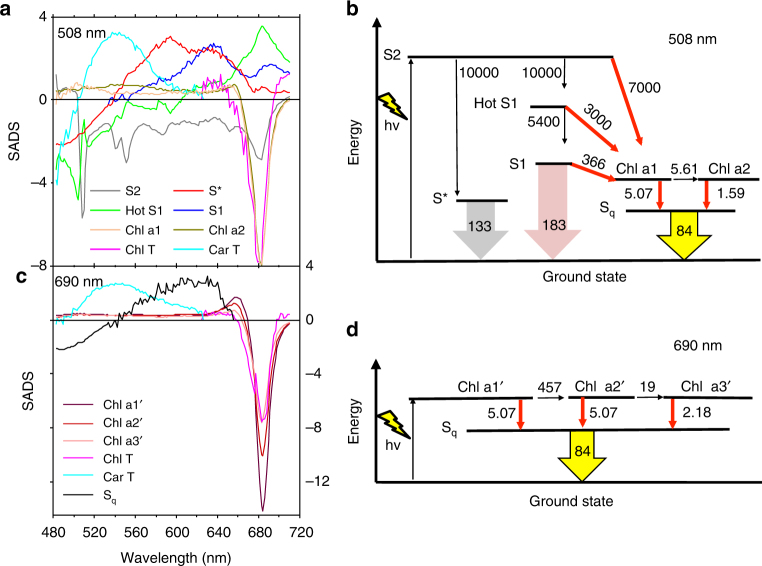
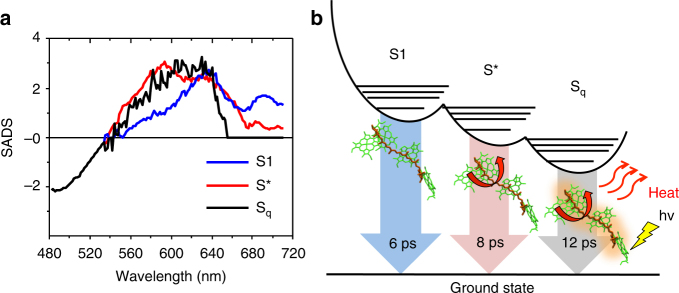
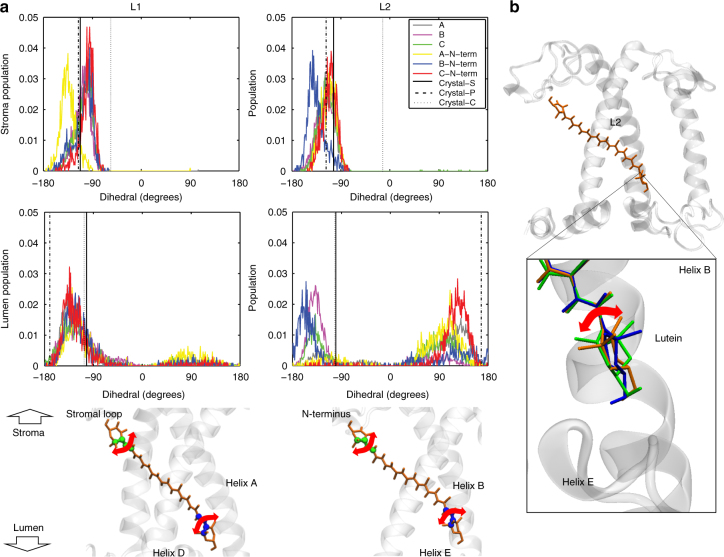
To avoid photodamage plants regulate the amount of excitation energy in the membrane at the level of the light-harvesting complexes (LHCs). It has been proposed that the energy absorbed in excess is dissipated via protein conformational changes of individual LHCs. However, the exact quenching mechanism remains unclear. Here we study the mechanism of quenching in LHCs that bind a single carotenoid species and are constitutively in a dissipative conformation. Via femtosecond spectroscopy we resolve a number of carotenoid dark states, demonstrating that the carotenoid is bound to the complex in different conformations. Some of those states act as excitation energy donors for the chlorophylls, whereas others act as quenchers. Via in silico analysis we show that structural changes of carotenoids are expected in the LHC protein domains exposed to the chloroplast lumen, where acidification triggers photoprotection in vivo. We propose that structural changes of LHCs control the conformation of the carotenoids, thus permitting access to different dark states responsible for either light harvesting or photoprotection.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Molecular factors controlling photosynthetic light harvesting by carotenoids.Acc Chem Res. 2010 Aug 17;43(8):1125-34. doi: 10.1021/ar100030m. Acc Chem Res. 2010. PMID: 20446691 Free PMC article.
-
Electron transfer between carotenoid and chlorophyll contributes to quenching in the LHCSR1 protein from Physcomitrella patens.Biochim Biophys Acta. 2016 Dec;1857(12):1870-1878. doi: 10.1016/j.bbabio.2016.09.001. Epub 2016 Sep 7. Biochim Biophys Acta. 2016. PMID: 27614061
-
Molecular adaptation of photoprotection: triplet states in light-harvesting proteins.Biophys J. 2011 Aug 17;101(4):934-42. doi: 10.1016/j.bpj.2011.05.057. Biophys J. 2011. PMID: 21843485 Free PMC article.
-
Quantum chemical insights in energy dissipation and carotenoid radical cation formation in light harvesting complexes.Phys Chem Chem Phys. 2007 Jun 21;9(23):2917-31. doi: 10.1039/b703028b. Epub 2007 Apr 25. Phys Chem Chem Phys. 2007. PMID: 17551615 Review.
-
Carotenoids and Photosynthesis.Subcell Biochem. 2016;79:111-39. doi: 10.1007/978-3-319-39126-7_4. Subcell Biochem. 2016. PMID: 27485220 Review.
Cited by
-
Light-harvesting complexes access analogue emissive states in different environments.Chem Sci. 2020 May 18;11(22):5697-5709. doi: 10.1039/d0sc00781a. eCollection 2020 Jun 14. Chem Sci. 2020. PMID: 32874506 Free PMC article.
-
Isomerization of carotenoids in photosynthesis and metabolic adaptation.Biophys Rev. 2023 Oct 11;15(5):887-906. doi: 10.1007/s12551-023-01156-4. eCollection 2023 Oct. Biophys Rev. 2023. PMID: 37974987 Free PMC article. Review.
-
Identification of distinct pH- and zeaxanthin-dependent quenching in LHCSR3 from Chlamydomonas reinhardtii.Elife. 2021 Jan 15;10:e60383. doi: 10.7554/eLife.60383. Elife. 2021. PMID: 33448262 Free PMC article.
-
Synthetic conversion of leaf chloroplasts into carotenoid-rich plastids reveals mechanistic basis of natural chromoplast development.Proc Natl Acad Sci U S A. 2020 Sep 1;117(35):21796-21803. doi: 10.1073/pnas.2004405117. Epub 2020 Aug 19. Proc Natl Acad Sci U S A. 2020. PMID: 32817419 Free PMC article.
-
The origin of pigment-binding differences in CP29 and LHCII: the role of protein structure and dynamics.Photochem Photobiol Sci. 2023 Jun;22(6):1279-1297. doi: 10.1007/s43630-023-00368-7. Epub 2023 Feb 6. Photochem Photobiol Sci. 2023. PMID: 36740636
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
