

Kinetics of drug-ribosome interactions defines the cidality of macrolide antibiotics
- PMID: 29229833
- PMCID: PMC5748224
- DOI: 10.1073/pnas.1717168115
Kinetics of drug-ribosome interactions defines the cidality of macrolide antibiotics
Abstract
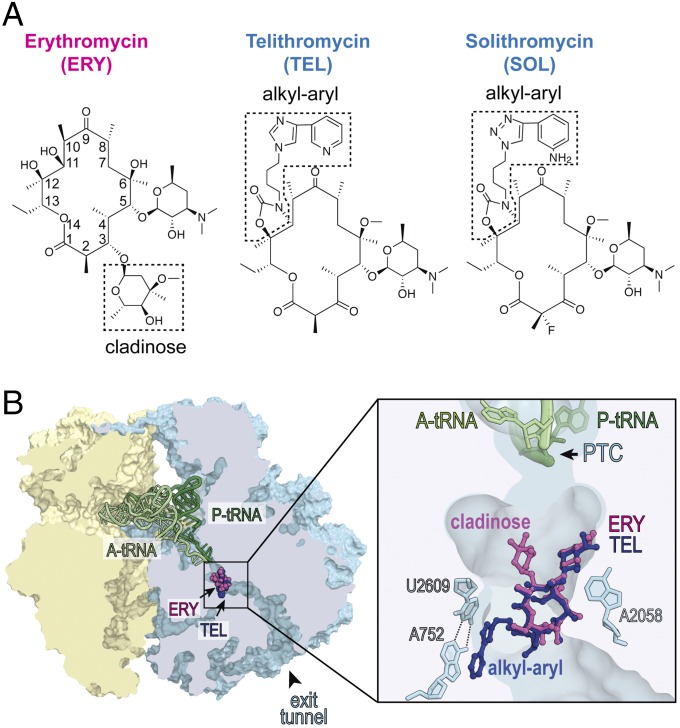
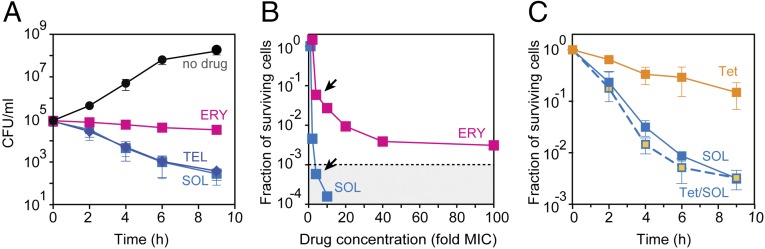
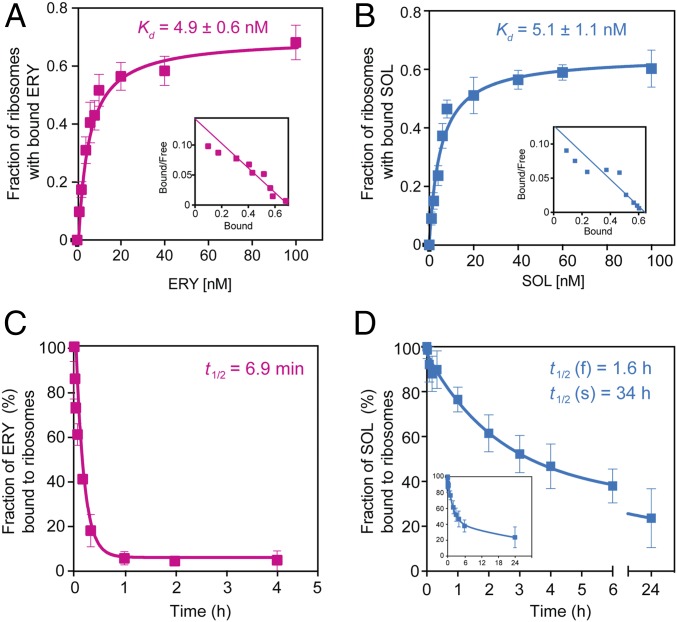
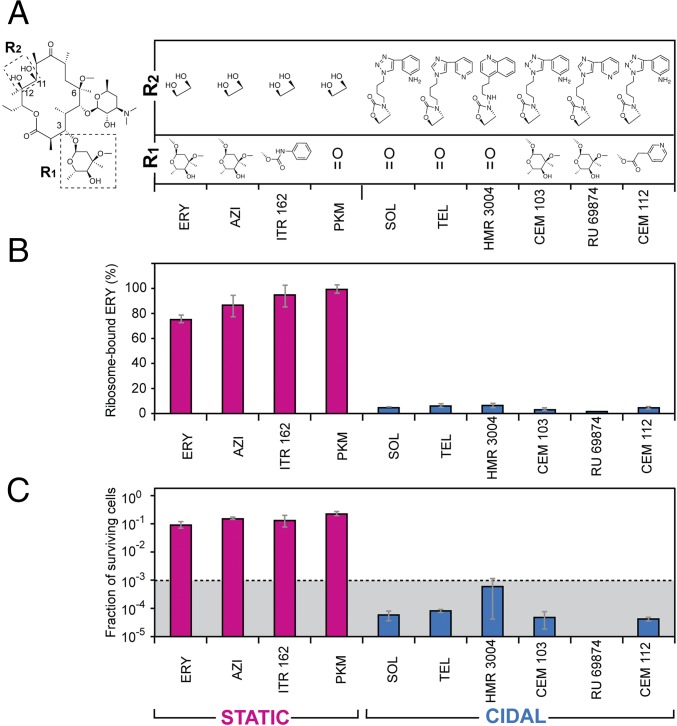
Antibiotics can cause dormancy (bacteriostasis) or induce death (cidality) of the targeted bacteria. The bactericidal capacity is one of the most important properties of antibacterial agents. However, the understanding of the fundamental differences in the mode of action of bacteriostatic or bactericidal antibiotics, especially those belonging to the same chemical class, is very rudimentary. Here, by examining the activity and binding properties of chemically distinct macrolide inhibitors of translation, we have identified a key difference in their interaction with the ribosome, which correlates with their ability to cause cell death. While bacteriostatic and bactericidal macrolides bind in the nascent peptide exit tunnel of the large ribosomal subunit with comparable affinities, the bactericidal antibiotics dissociate from the ribosome with significantly slower rates. The sluggish dissociation of bactericidal macrolides correlates with the presence in their structure of an extended alkyl-aryl side chain, which establishes idiosyncratic interactions with the ribosomal RNA. Mutations or chemical alterations of the rRNA nucleotides in the drug binding site can protect cells from macrolide-induced killing, even with inhibitor concentrations that significantly exceed those required for cell growth arrest. We propose that the increased translation downtime due to slow dissociation of the antibiotic may damage cells beyond the point where growth can be reinitiated upon the removal of the drug due to depletion of critical components of the gene-expression pathway.
Keywords: erythromycin; inhibitors; ketolides; solithromycin; translation.
Conflict of interest statement
Conflict of interest statement: In the previous years, A.S.M. had grants from Cempra, Inc. and Melinta Therapeutics, which were involved in the development of macrolide antibiotics.
Figures
References
-
- Clinical and Laboratory Standards Institute . Methods for Determining Bactericidal Activity of Antimicrobial Agents; Approved Guideline. NCCLS Document M26-A. NCCLS; Wayne, PA: 1999.
-
- Goldstein FW, Emirian MF, Coutrot A, Acar JF. Bacteriostatic and bactericidal activity of azithromycin against Haemophilus influenzae. J Antimicrob Chemother. 1990;25(Suppl A):25–28. - PubMed
-
- Gaynor M, Mankin AS. Macrolide antibiotics: Binding site, mechanism of action, resistance. Curr Top Med Chem. 2003;3:949–961. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
