

Myosin-X knockout is semi-lethal and demonstrates that myosin-X functions in neural tube closure, pigmentation, hyaloid vasculature regression, and filopodia formation
- PMID: 29229982
- PMCID: PMC5725431
- DOI: 10.1038/s41598-017-17638-x
Myosin-X knockout is semi-lethal and demonstrates that myosin-X functions in neural tube closure, pigmentation, hyaloid vasculature regression, and filopodia formation
Abstract
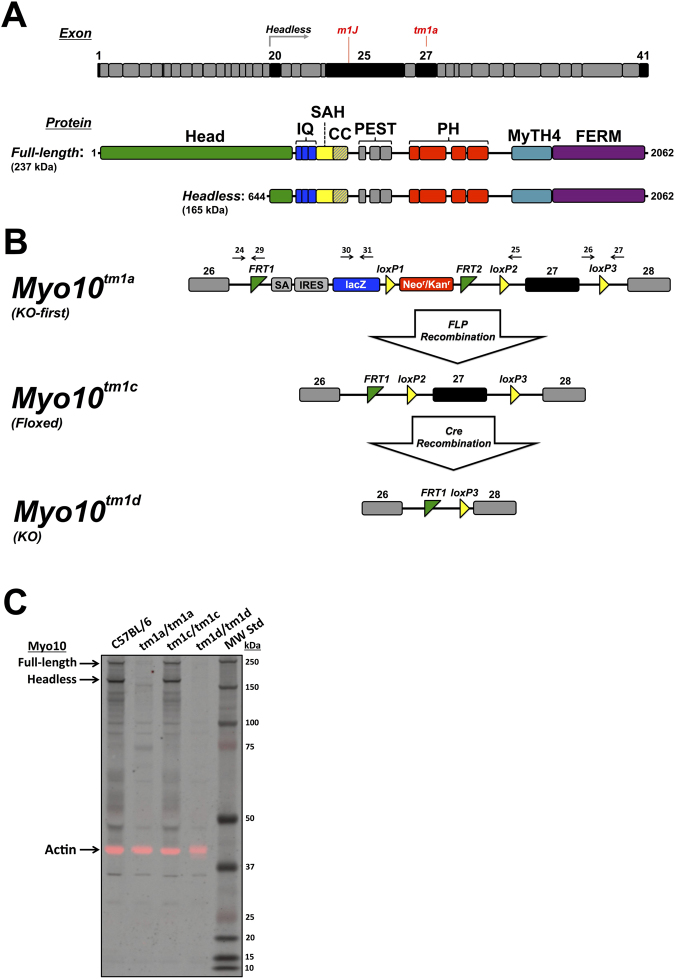
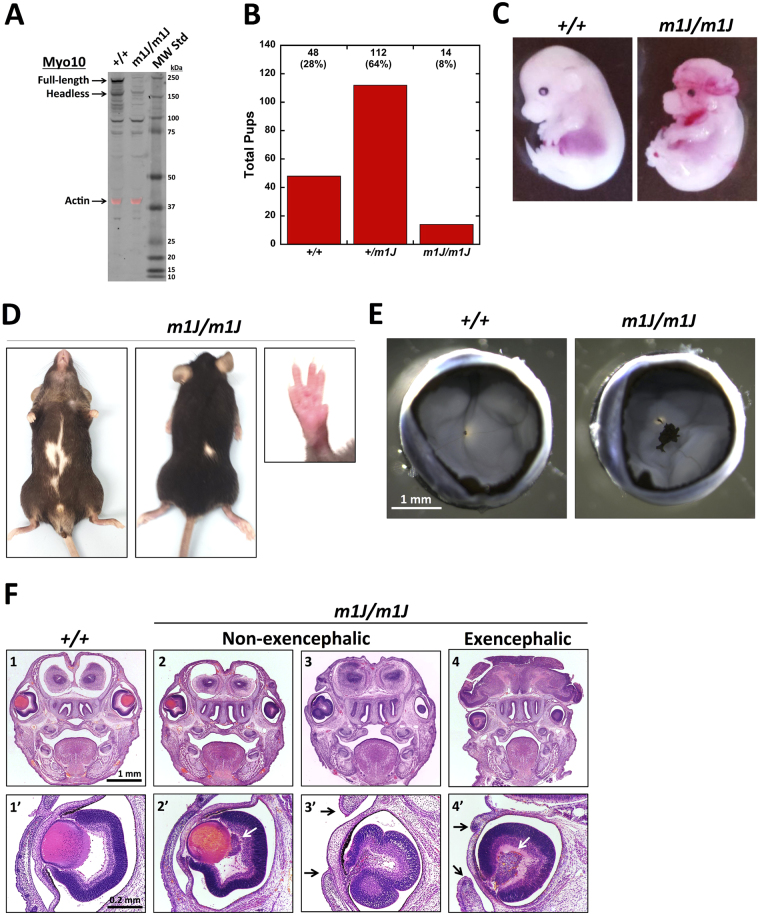
Myosin-X (Myo10) is an unconventional myosin best known for its striking localization to the tips of filopodia. Despite the broad expression of Myo10 in vertebrate tissues, its functions at the organismal level remain largely unknown. We report here the generation of KO-first (Myo10 tm1a/tm1a ), floxed (Myo10 tm1c/tm1c ), and KO mice (Myo10 tm1d/tm1d ). Complete knockout of Myo10 is semi-lethal, with over half of homozygous KO embryos exhibiting exencephaly, a severe defect in neural tube closure. All Myo10 KO mice that survive birth exhibit a white belly spot, all have persistent fetal vasculature in the eye, and ~50% have webbed digits. Myo10 KO mice that survive birth can breed and produce litters of KO embryos, demonstrating that Myo10 is not absolutely essential for mitosis, meiosis, adult survival, or fertility. KO-first mice and an independent spontaneous deletion (Myo10 m1J/m1J ) exhibit the same core phenotypes. During retinal angiogenesis, KO mice exhibit a ~50% decrease in endothelial filopodia, demonstrating that Myo10 is required to form normal numbers of filopodia in vivo. The Myo10 mice generated here demonstrate that Myo10 has important functions in mammalian development and provide key tools for defining the functions of Myo10 in vivo.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
-
- Berg JS, Derfler BH, Pennisi CM, Corey DP, Cheney RE. Myosin-X, a novel myosin with pleckstrin homology domains, associates with regions of dynamic actin. J Cell Sci. 2000;113:3439–3451. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
