

Structure-based membrane dome mechanism for Piezo mechanosensitivity
- PMID: 29231809
- PMCID: PMC5788504
- DOI: 10.7554/eLife.33660
Structure-based membrane dome mechanism for Piezo mechanosensitivity
Abstract
Mechanosensitive ion channels convert external mechanical stimuli into electrochemical signals for critical processes including touch sensation, balance, and cardiovascular regulation. The best understood mechanosensitive channel, MscL, opens a wide pore, which accounts for mechanosensitive gating due to in-plane area expansion. Eukaryotic Piezo channels have a narrow pore and therefore must capture mechanical forces to control gating in another way. We present a cryo-EM structure of mouse Piezo1 in a closed conformation at 3.7Å-resolution. The channel is a triskelion with arms consisting of repeated arrays of 4-TM structural units surrounding a pore. Its shape deforms the membrane locally into a dome. We present a hypothesis in which the membrane deformation changes upon channel opening. Quantitatively, membrane tension will alter gating energetics in proportion to the change in projected area under the dome. This mechanism can account for highly sensitive mechanical gating in the setting of a narrow, cation-selective pore.
Keywords: Piezo channel; biophysics; cryoEM; mechanosensitivity; mouse; structural biology.
© 2017, Guo et al.
Conflict of interest statement
YG, RM No competing interests declared
Figures








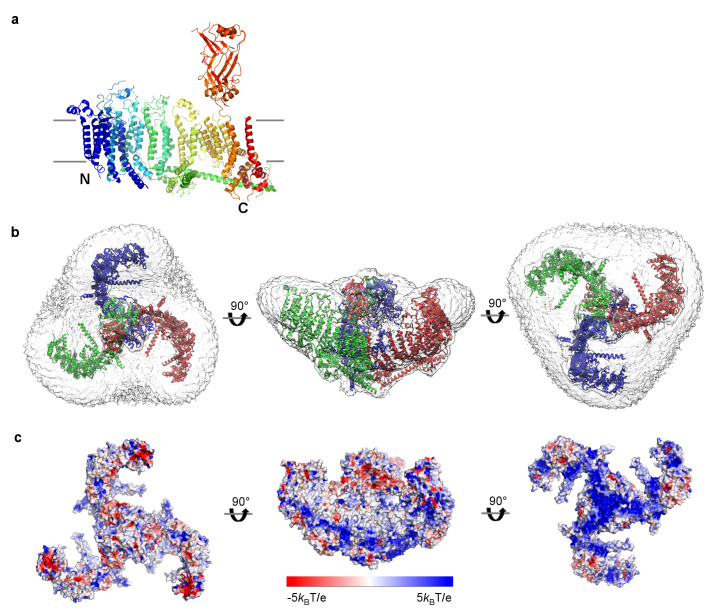



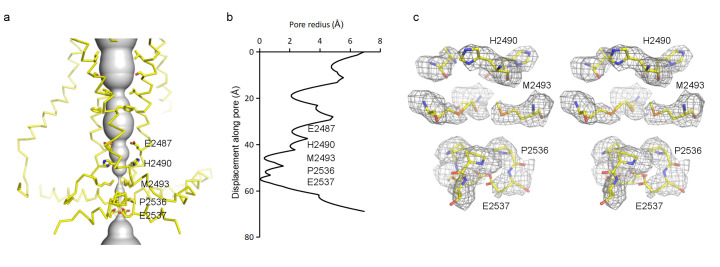



Comment in
-
Portraits of a pressure sensor.Elife. 2018 Jan 29;7:e34396. doi: 10.7554/eLife.34396. Elife. 2018. PMID: 29376828 Free PMC article.
References
-
- Afonine P, Headd J, Terwilliger T, Adams P. New tool: Phenix.real-space-refine. Computational Crystallography Newsletter. 2013;4:43–44.
-
- Alper SL. Current Topics in Membranes. Elsevier; 2017. Genetic Diseases of PIEZO1 and PIEZO2 Dysfunction. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
