

Large Cardiac Muscle Patches Engineered From Human Induced-Pluripotent Stem Cell-Derived Cardiac Cells Improve Recovery From Myocardial Infarction in Swine
- PMID: 29233823
- PMCID: PMC5903991
- DOI: 10.1161/CIRCULATIONAHA.117.030785
Large Cardiac Muscle Patches Engineered From Human Induced-Pluripotent Stem Cell-Derived Cardiac Cells Improve Recovery From Myocardial Infarction in Swine
Abstract
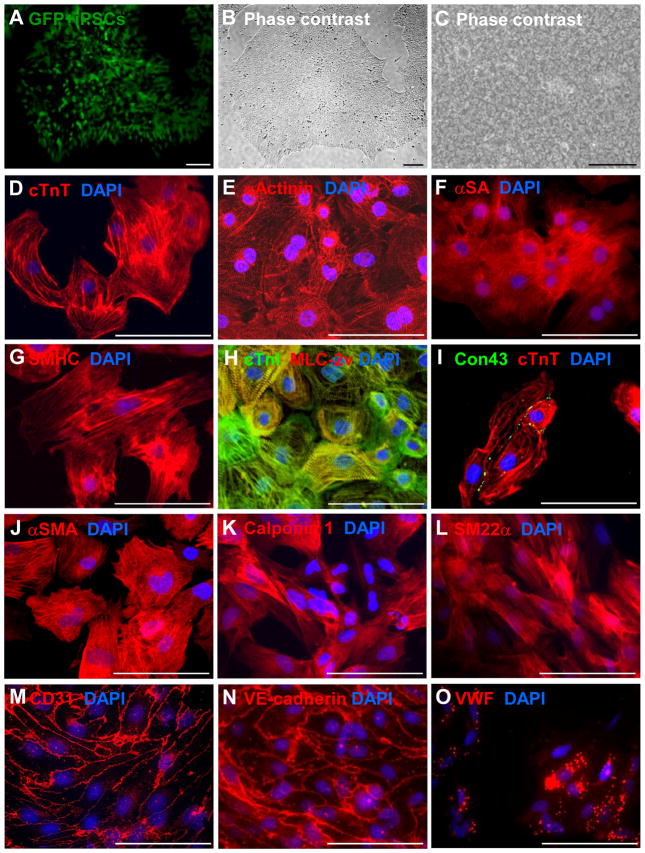
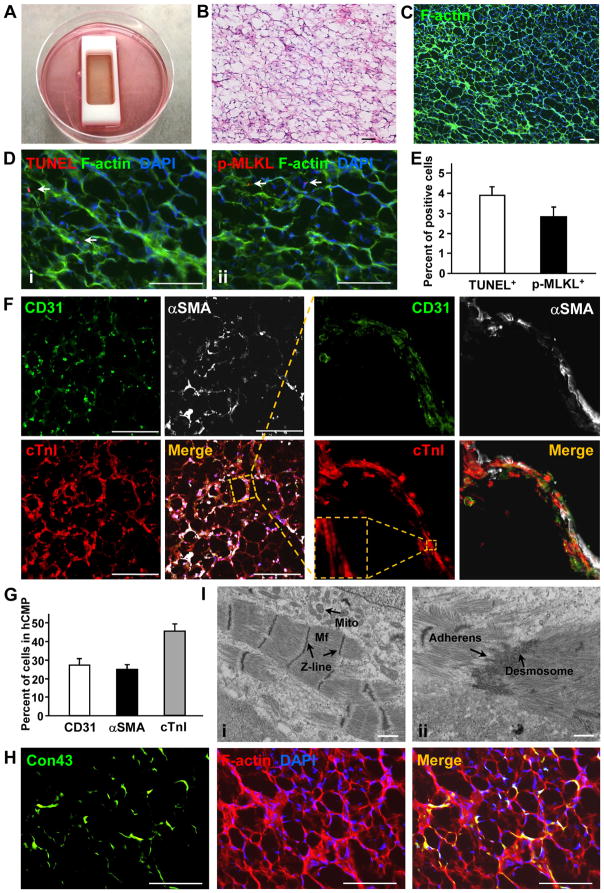
Background: Here, we generated human cardiac muscle patches (hCMPs) of clinically relevant dimensions (4 cm × 2 cm × 1.25 mm) by suspending cardiomyocytes, smooth muscle cells, and endothelial cells that had been differentiated from human induced-pluripotent stem cells in a fibrin scaffold and then culturing the construct on a dynamic (rocking) platform.
Methods: In vitro assessments of hCMPs suggest maturation in response to dynamic culture stimulation. In vivo assessments were conducted in a porcine model of myocardial infarction (MI). Animal groups included: MI hearts treated with 2 hCMPs (MI+hCMP, n=13), MI hearts treated with 2 cell-free open fibrin patches (n=14), or MI hearts with neither experimental patch (n=15); a fourth group of animals underwent sham surgery (Sham, n=8). Cardiac function and infarct size were evaluated by MRI, arrhythmia incidence by implanted loop recorders, and the engraftment rate by calculation of quantitative polymerase chain reaction measurements of expression of the human Y chromosome. Additional studies examined the myocardial protein expression profile changes and potential mechanisms of action that related to exosomes from the cell patch.
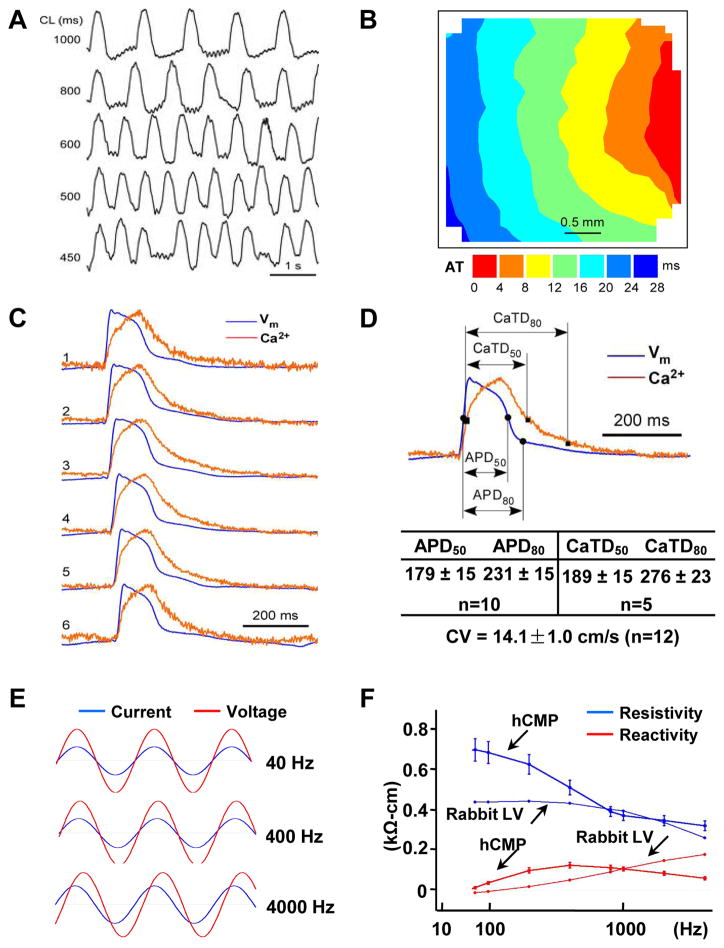
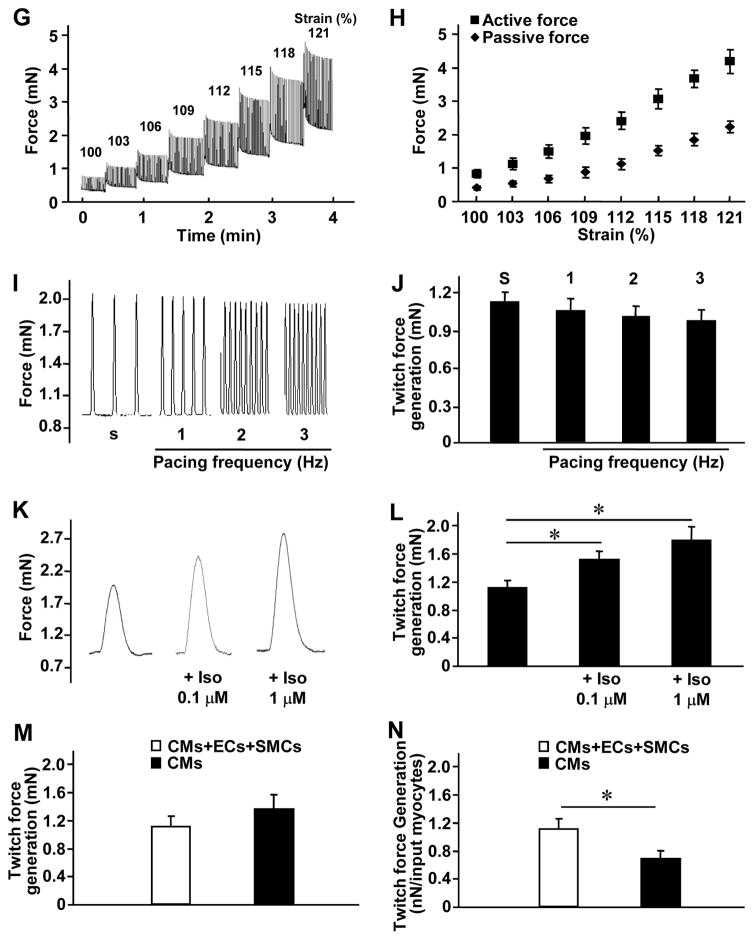
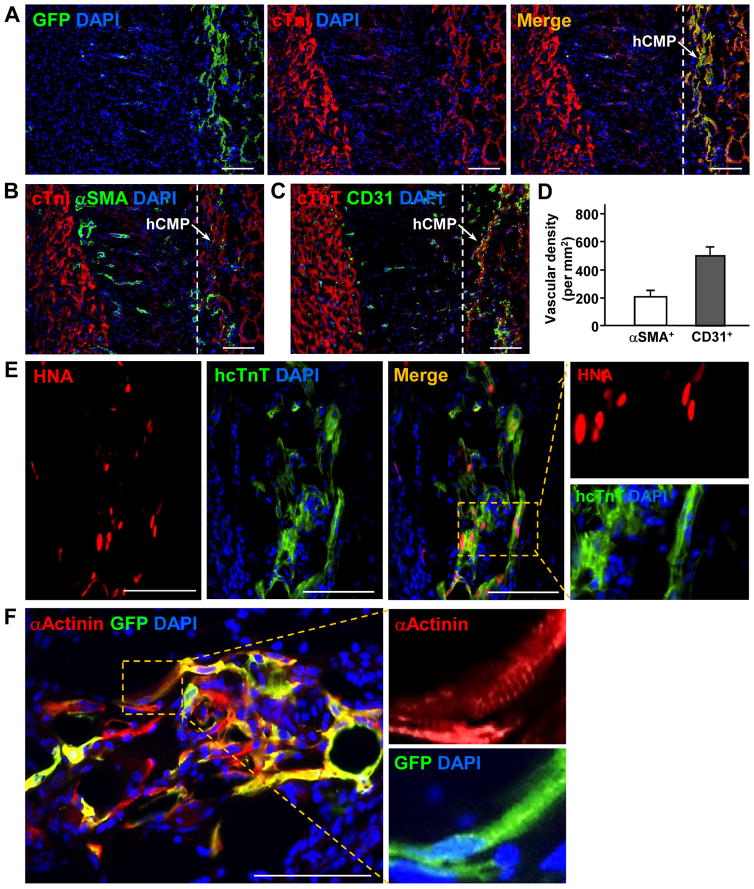
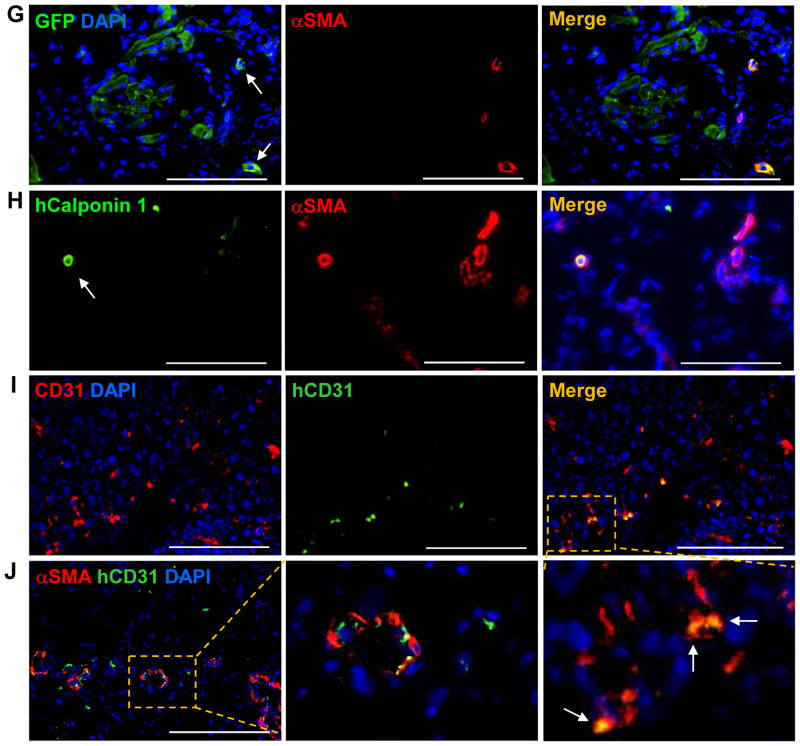
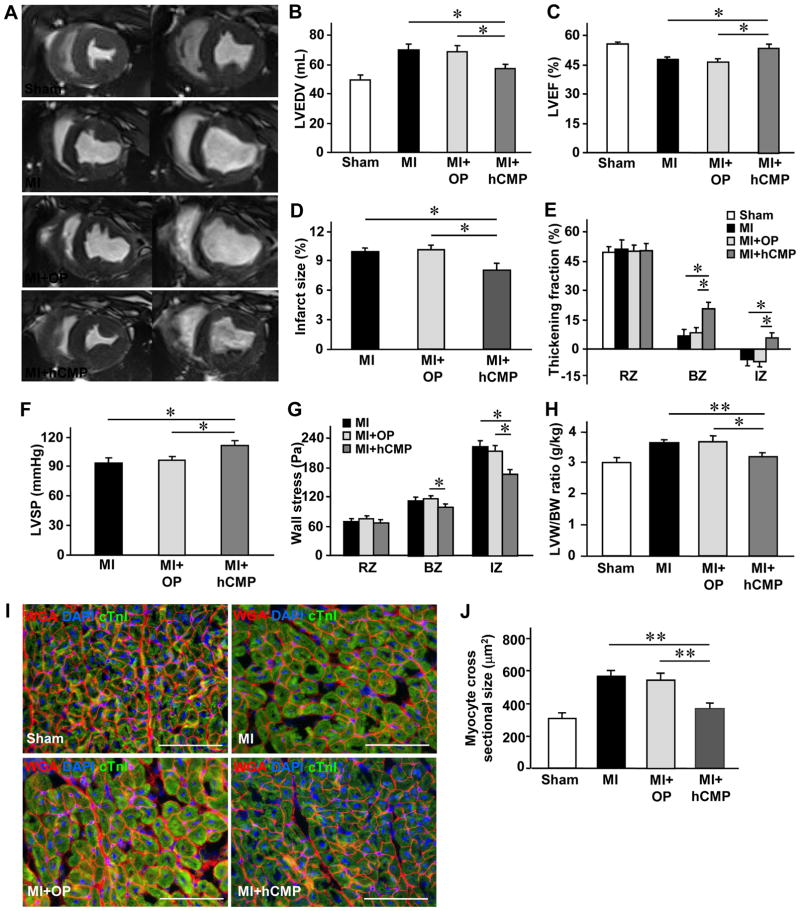
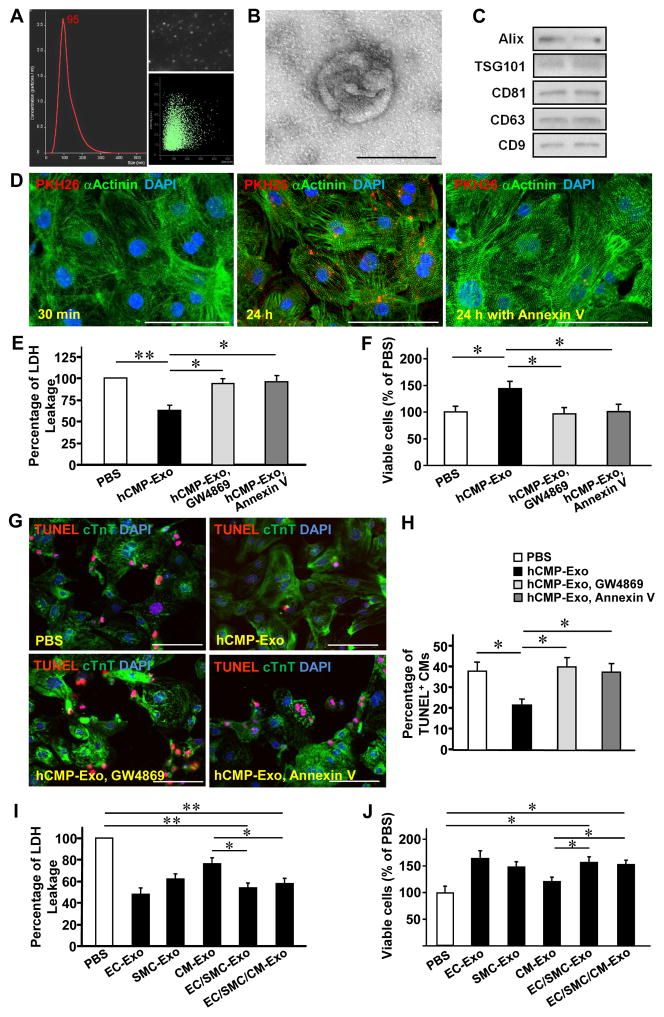
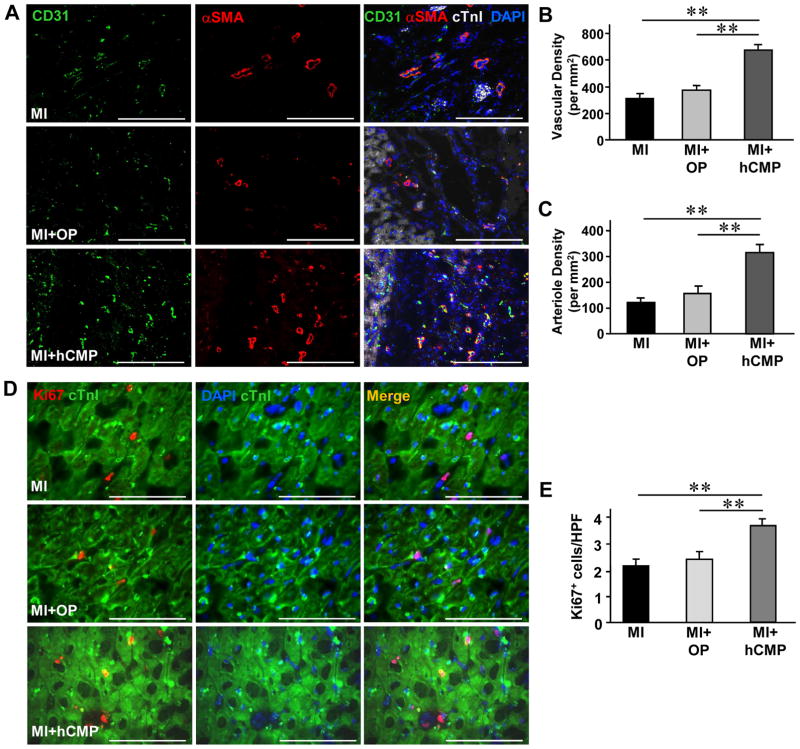
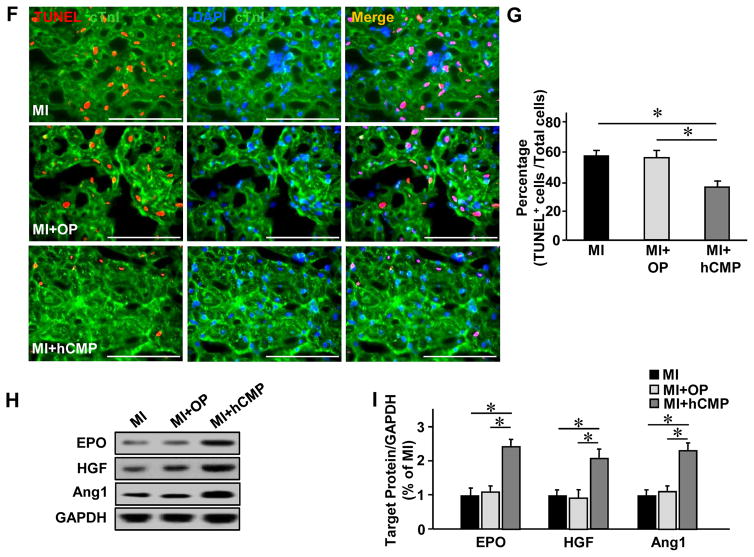
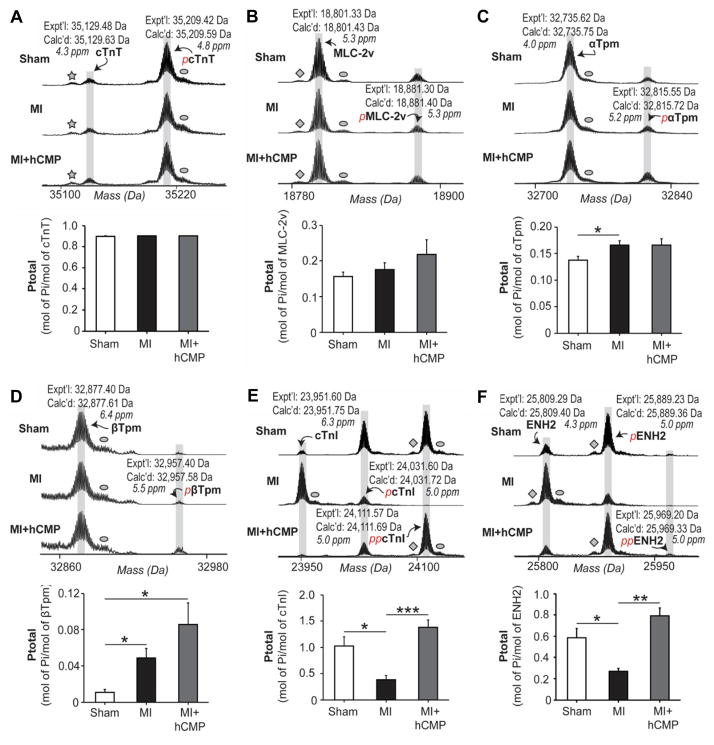
Results: The hCMPs began to beat synchronously within 1 day of fabrication, and after 7 days of dynamic culture stimulation, in vitro assessments indicated the mechanisms related to the improvements in electronic mechanical coupling, calcium-handling, and force generation, suggesting a maturation process during the dynamic culture. The engraftment rate was 10.9±1.8% at 4 weeks after the transplantation. The hCMP transplantation was associated with significant improvements in left ventricular function, infarct size, myocardial wall stress, myocardial hypertrophy, and reduced apoptosis in the periscar boarder zone myocardium. hCMP transplantation also reversed some MI-associated changes in sarcomeric regulatory protein phosphorylation. The exosomes released from the hCMP appeared to have cytoprotective properties that improved cardiomyocyte survival.
Conclusions: We have fabricated a clinically relevant size of hCMP with trilineage cardiac cells derived from human induced-pluripotent stem cells. The hCMP matures in vitro during 7 days of dynamic culture. Transplantation of this type of hCMP results in significantly reduced infarct size and improvements in cardiac function that are associated with reduction in left ventricular wall stress. The hCMP treatment is not associated with significant changes in arrhythmogenicity.
Keywords: heart; models, animal; myocardial infarction; pluripotent stem cells; tissue engineering.
© 2017 American Heart Association, Inc.
Figures
References
-
- Shiba Y, Gomibuchi T, Seto T, Wada Y, Ichimura H, Tanaka Y, Ogasawara T, Okada K, Shiba N, Sakamoto K, Ido D, Shiina T, Ohkura M, Nakai J, Uno N, Kazuki Y, Oshimura M, Minami I, Ikeda U. Allogeneic transplantation of iPS cell-derived cardiomyocytes regenerates primate hearts. Nature. 2016;538:388–391. doi: 10.1038/nature19815. - DOI - PubMed
-
- Ye L, Chang YH, Xiong Q, Zhang P, Zhang L, Somasundaram P, Lepley M, Swingen C, Su L, Wendel JS, Guo J, Jang A, Rosenbush D, Greder L, Dutton JR, Zhang J, Kamp TJ, Kaufman DS, Ge Y, Zhang J. Cardiac repair in a porcine model of acute myocardial infarction with human induced pluripotent stem cell-derived cardiovascular cells. Cell Stem Cell. 2014;15:750–61. doi: 10.1016/j.stem.2014.11.009. - DOI - PMC - PubMed
-
- Riegler J, Tiburcy M, Ebert A, Tzatzalos E, Raaz U, Abilez OJ, Shen Q, Kooreman NG, Neofytou E, Chen VC, Wang M, Meyer T, Tsao PS, Connolly AJ, Couture LA, Gold JD, Zimmermann WH, Wu JC. Human Engineered Heart Muscles Engraft and Survive Long Term in a Rodent Myocardial Infarction Model. Circulation research. 2015;117:720–30. doi: 10.1161/CIRCRESAHA.115.306985. - DOI - PMC - PubMed
-
- Chong JJ, Yang X, Don CW, Minami E, Liu YW, Weyers JJ, Mahoney WM, Van Biber B, Cook SM, Palpant NJ, Gantz JA, Fugate JA, Muskheli V, Gough GM, Vogel KW, Astley CA, Hotchkiss CE, Baldessari A, Pabon L, Reinecke H, Gill EA, Nelson V, Kiem HP, Laflamme MA, Murry CE. Human embryonic-stem-cell-derived cardiomyocytes regenerate non-human primate hearts. Nature. 2014;510:273–7. doi: 10.1038/nature13233. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
