

A perspective on Na and K channel inactivation
- PMID: 29233885
- PMCID: PMC5749110
- DOI: 10.1085/jgp.201711835
A perspective on Na and K channel inactivation
Abstract
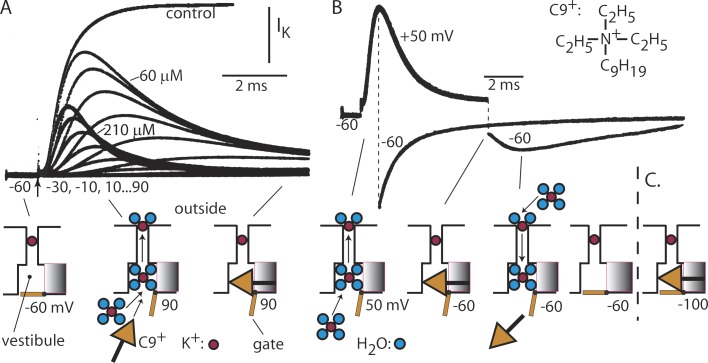
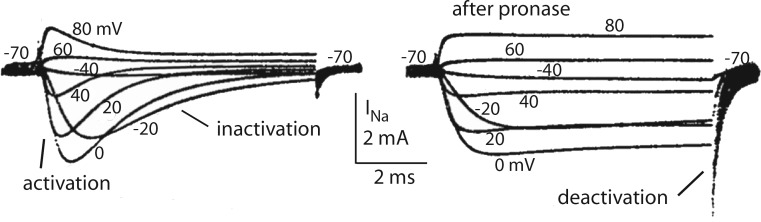
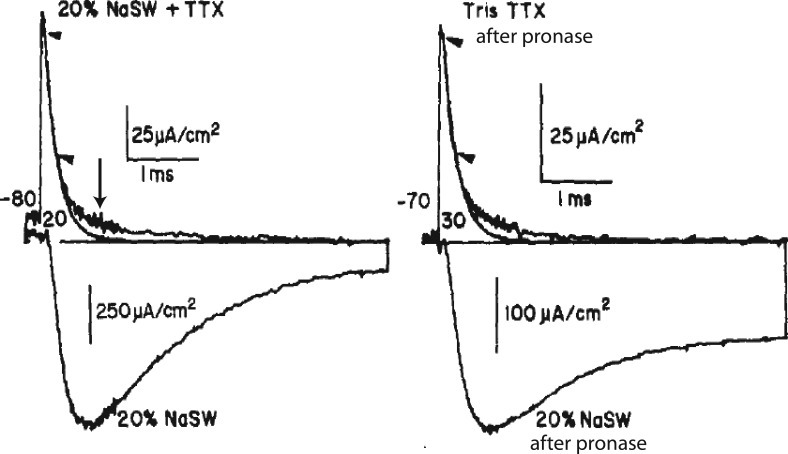
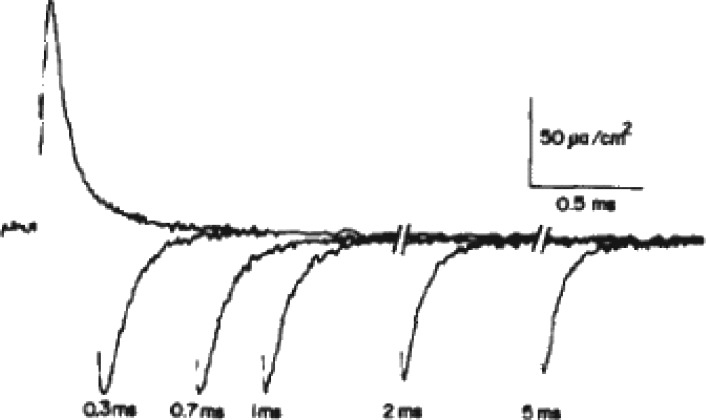
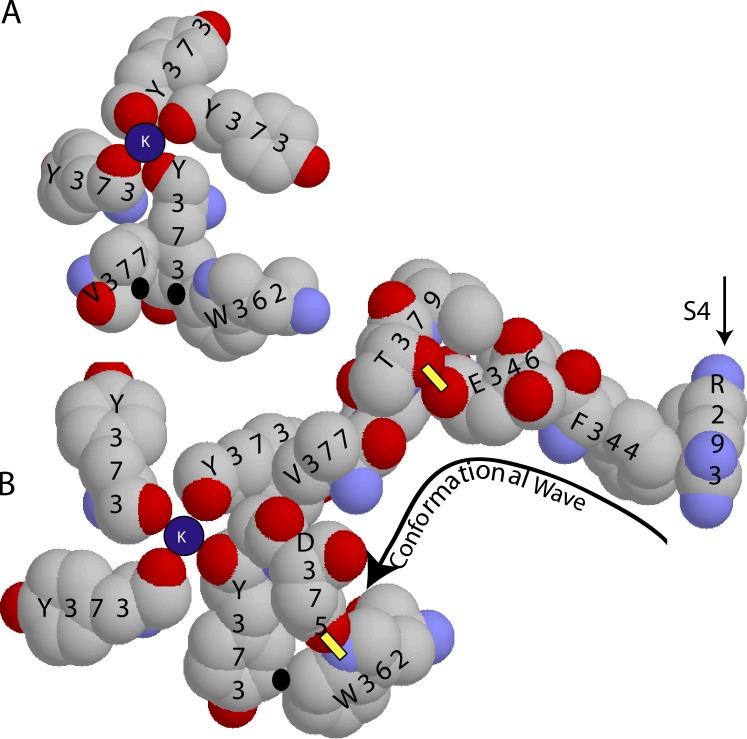
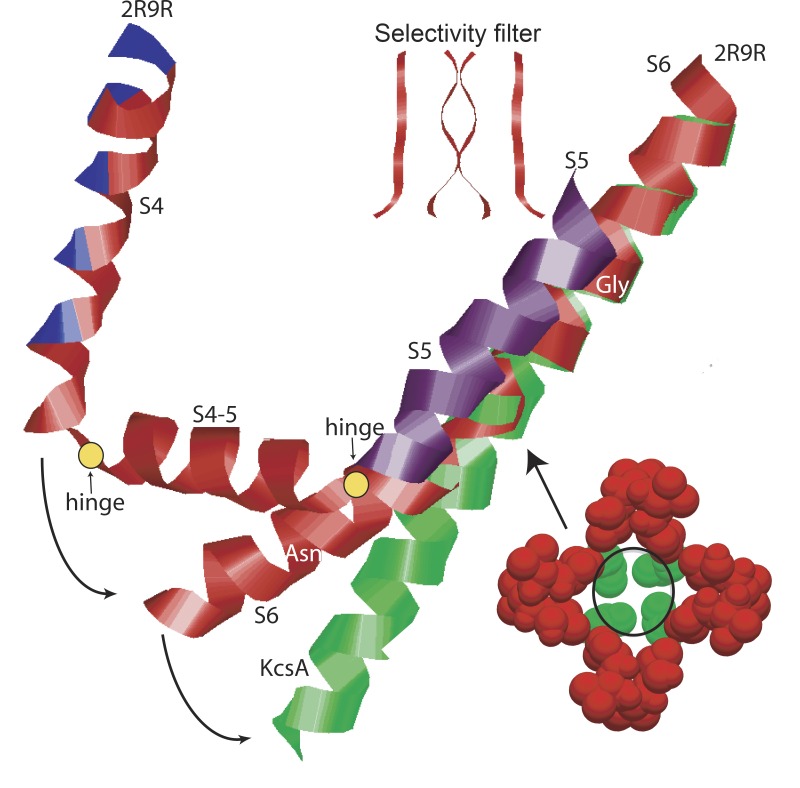
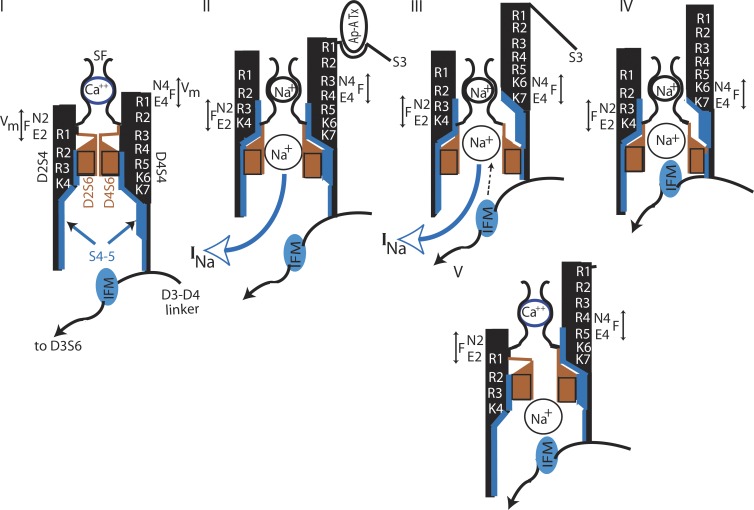
We are wired with conducting cables called axons that rapidly transmit electrical signals (e.g., "Ouch!") from, for example, the toe to the spinal cord. Because of the high internal resistance of axons (salt water rather than copper), a signal must be reinforced after traveling a short distance. Reinforcement is accomplished by ion channels, Na channels for detecting the signal and reinforcing it by driving it further positive (to near 50 mV) and K channels for then restoring it to the resting level (near -70 mV). The signal is called an action potential and has a duration of roughly a millisecond. The return of membrane voltage (Vm) to the resting level after an action potential is facilitated by "inactivation" of the Na channels: i.e., an internal particle diffuses into the mouth of any open Na channel and temporarily blocks it. Some types of K channels also show inactivation after being open for a time. N-type inactivation of K channels has a relatively fast time course and involves diffusion of the N-terminal of one of the channel's four identical subunits into the channel's inner mouth, if it is open. This mechanism is similar to Na channel inactivation. Both Na and K channels also display slower inactivation processes. C inactivation in K channels involves changes in the channel's outer mouth, the "selectivity filter," whose normal function is to prevent Na+ ions from entering the K channel. C inactivation deforms the filter so that neither K+ nor Na+ can pass.
© 2018 Armstrong and Hollingworth.
Figures
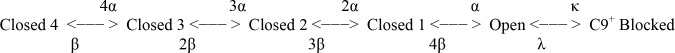
References
-
- Armstrong C.M. 1981. Sodium channels and gating currents. Physiol. Rev. 61:644–683. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
