

Prophages and Growth Dynamics Confound Experimental Results with Antibiotic-Tolerant Persister Cells
- PMID: 29233898
- PMCID: PMC5727415
- DOI: 10.1128/mBio.01964-17
Prophages and Growth Dynamics Confound Experimental Results with Antibiotic-Tolerant Persister Cells
Abstract
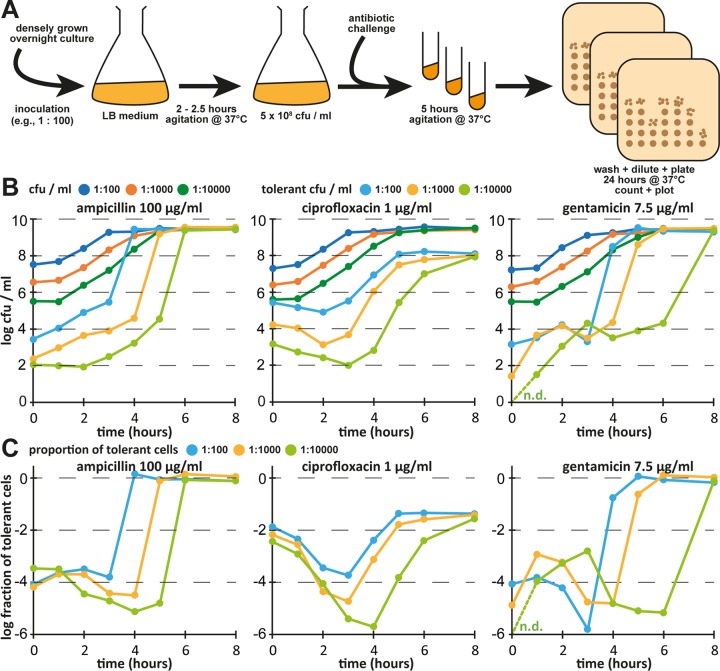
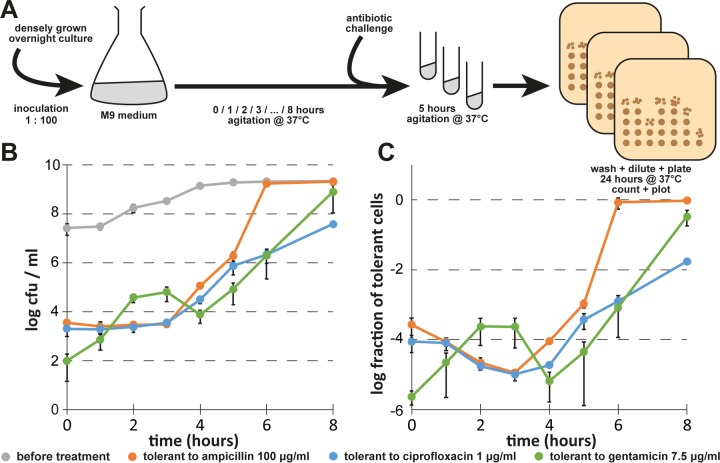
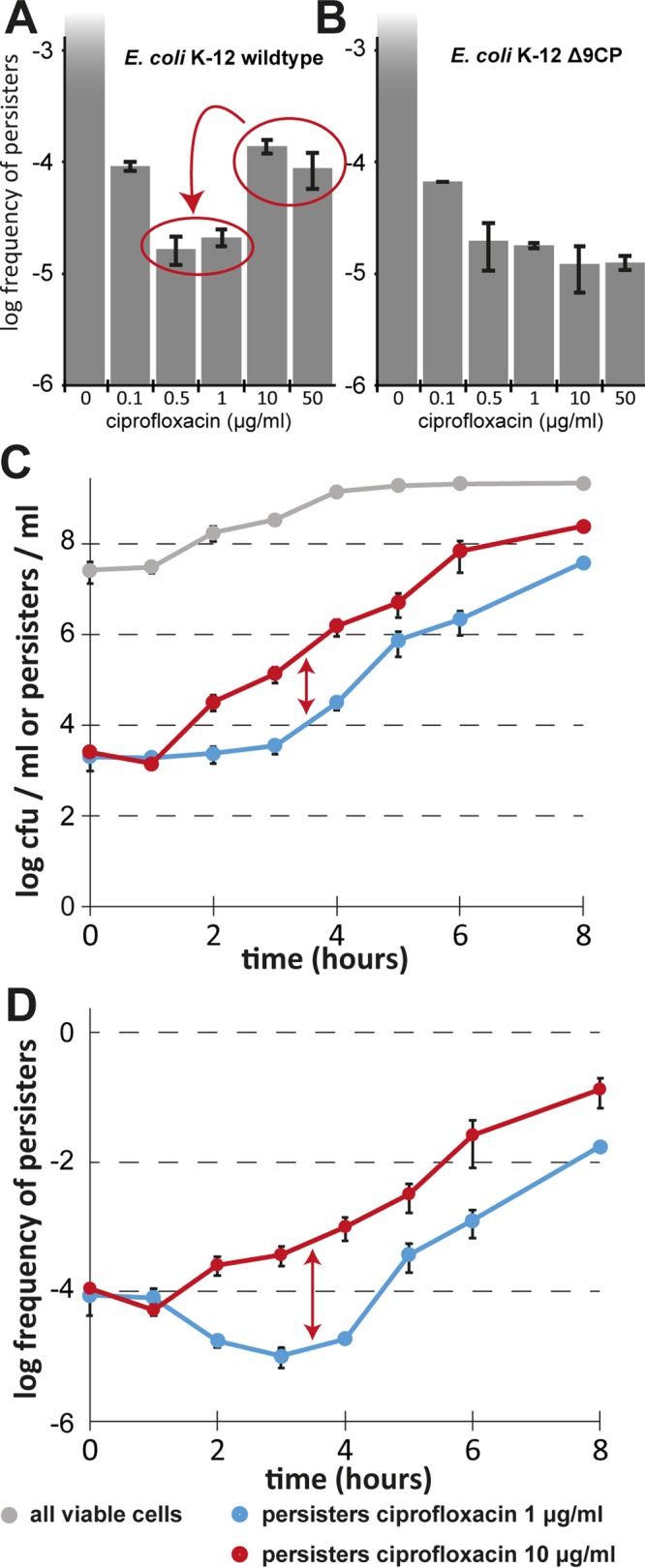
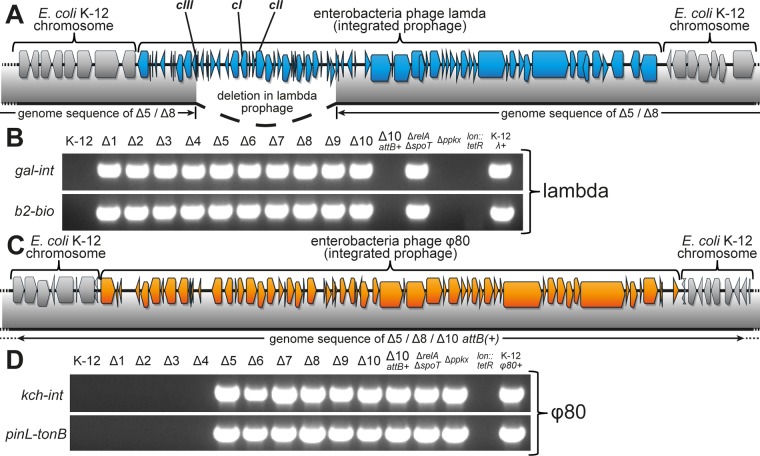
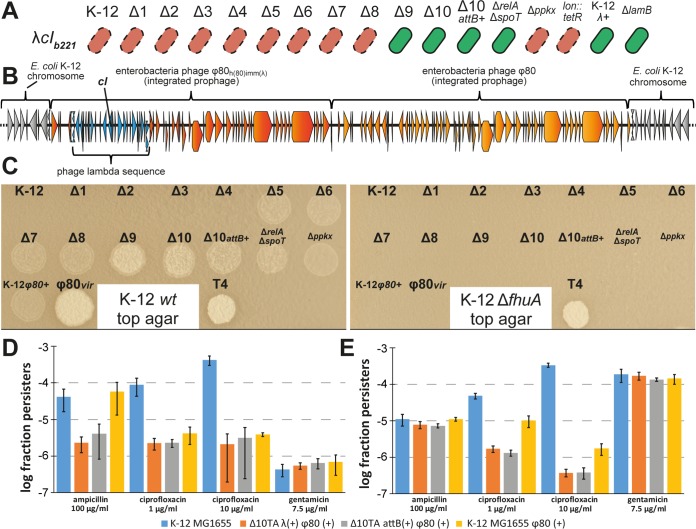
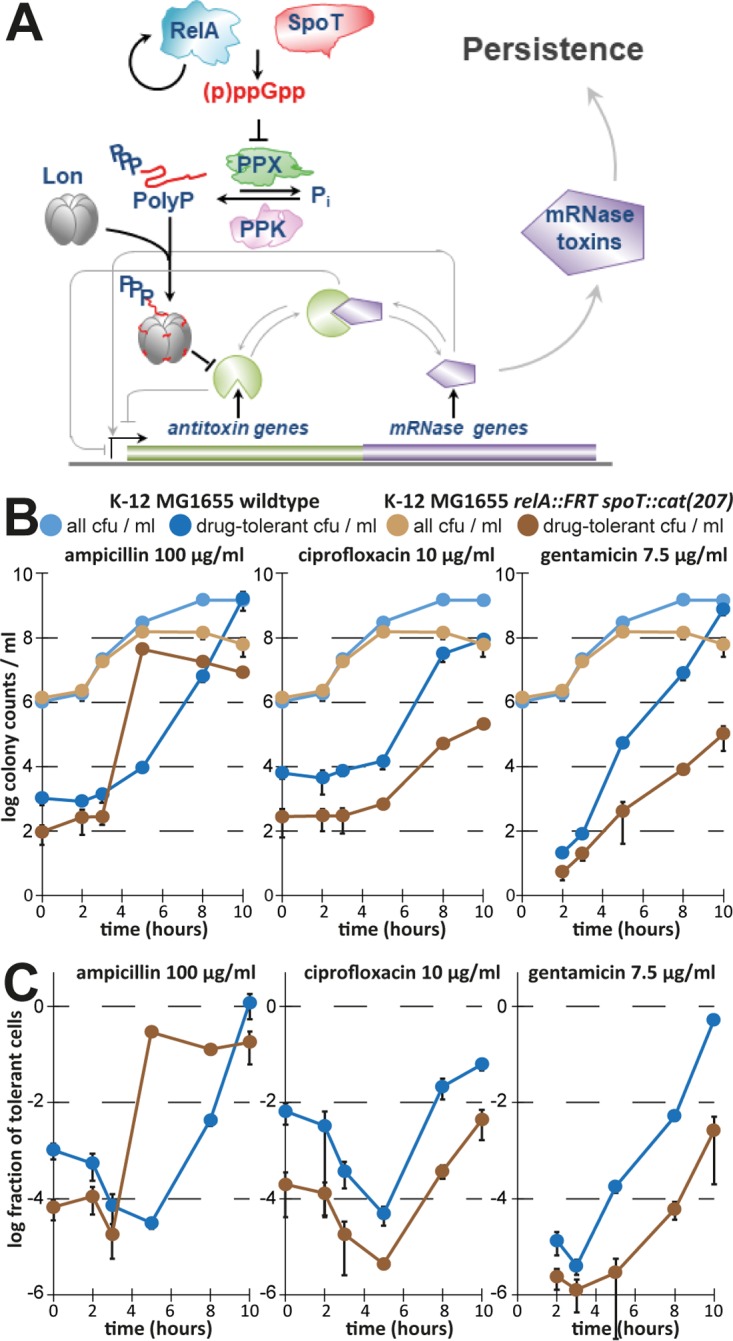
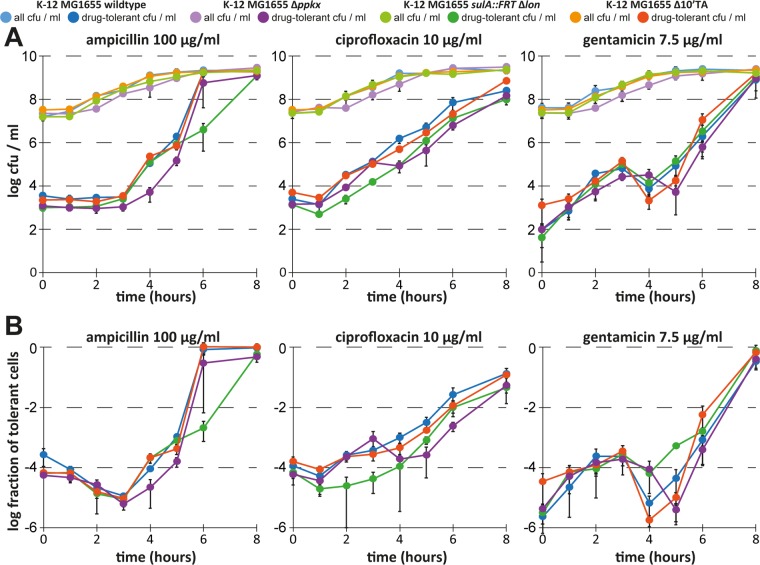
Bacterial persisters are phenotypic variants that survive antibiotic treatment in a dormant state and can be formed by multiple pathways. We recently proposed that the second messenger (p)ppGpp drives Escherichia coli persister formation through protease Lon and activation of toxin-antitoxin (TA) modules. This model found considerable support among researchers studying persisters but also generated controversy as part of recent debates in the field. In this study, we therefore used our previous work as a model to critically examine common experimental procedures to understand and overcome the inconsistencies often observed between results of different laboratories. Our results show that seemingly simple antibiotic killing assays are very sensitive to variations in culture conditions and bacterial growth phase. Additionally, we found that some assay conditions cause the killing of antibiotic-tolerant persisters via induction of cryptic prophages. Similarly, the inadvertent infection of mutant strains with bacteriophage ϕ80, a notorious laboratory contaminant, apparently caused several of the phenotypes that we reported in our previous studies. We therefore reconstructed all infected mutants and probed the validity of our model of persister formation in a refined assay setup that uses robust culture conditions and unravels the dynamics of persister cells through all bacterial growth stages. Our results confirm the importance of (p)ppGpp and Lon but no longer support a role of TA modules in E. coli persister formation under unstressed conditions. We anticipate that the results and approaches reported in our study will lay the ground for future work in the field.IMPORTANCE The recalcitrance of antibiotic-tolerant persister cells is thought to cause relapsing infections and antibiotic treatment failure in various clinical setups. Previous studies identified multiple genetic pathways involved in persister formation but also revealed reproducibility problems that sparked controversies about adequate tools to study persister cells. In this study, we unraveled how typical antibiotic killing assays often fail to capture the biology of persisters and instead give widely differing results based on poorly controlled experimental parameters and artifacts caused by cryptic as well as contaminant prophages. We therefore established a new, robust assay that enabled us to follow the dynamics of persister cells through all growth stages of bacterial cultures without distortions by bacteriophages. This system also favored adequate comparisons of mutant strains with aberrant growth phenotypes. We anticipate that our results will contribute to a robust, common basis for future studies on the formation and eradication of antibiotic-tolerant persisters.
Keywords: (p)ppGpp; antibiotic tolerance; bacteriophage genetics; persistence; toxin-antitoxin modules.
Copyright © 2017 Harms et al.
Figures
References
-
- Verstraeten N, Knapen WJ, Kint CI, Liebens V, Van den Bergh B, Dewachter L, Michiels JE, Fu Q, David CC, Fierro AC, Marchal K, Beirlant J, Versées W, Hofkens J, Jansen M, Fauvart M, Michiels J. 2015. Obg and membrane depolarization are part of a microbial bet-hedging strategy that leads to antibiotic tolerance. Mol Cell 59:9–21. doi: 10.1016/j.molcel.2015.05.011. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
