

ATG-dependent phagocytosis in dendritic cells drives myelin-specific CD4+ T cell pathogenicity during CNS inflammation
- PMID: 29233943
- PMCID: PMC5748192
- DOI: 10.1073/pnas.1713664114
ATG-dependent phagocytosis in dendritic cells drives myelin-specific CD4+ T cell pathogenicity during CNS inflammation
Abstract
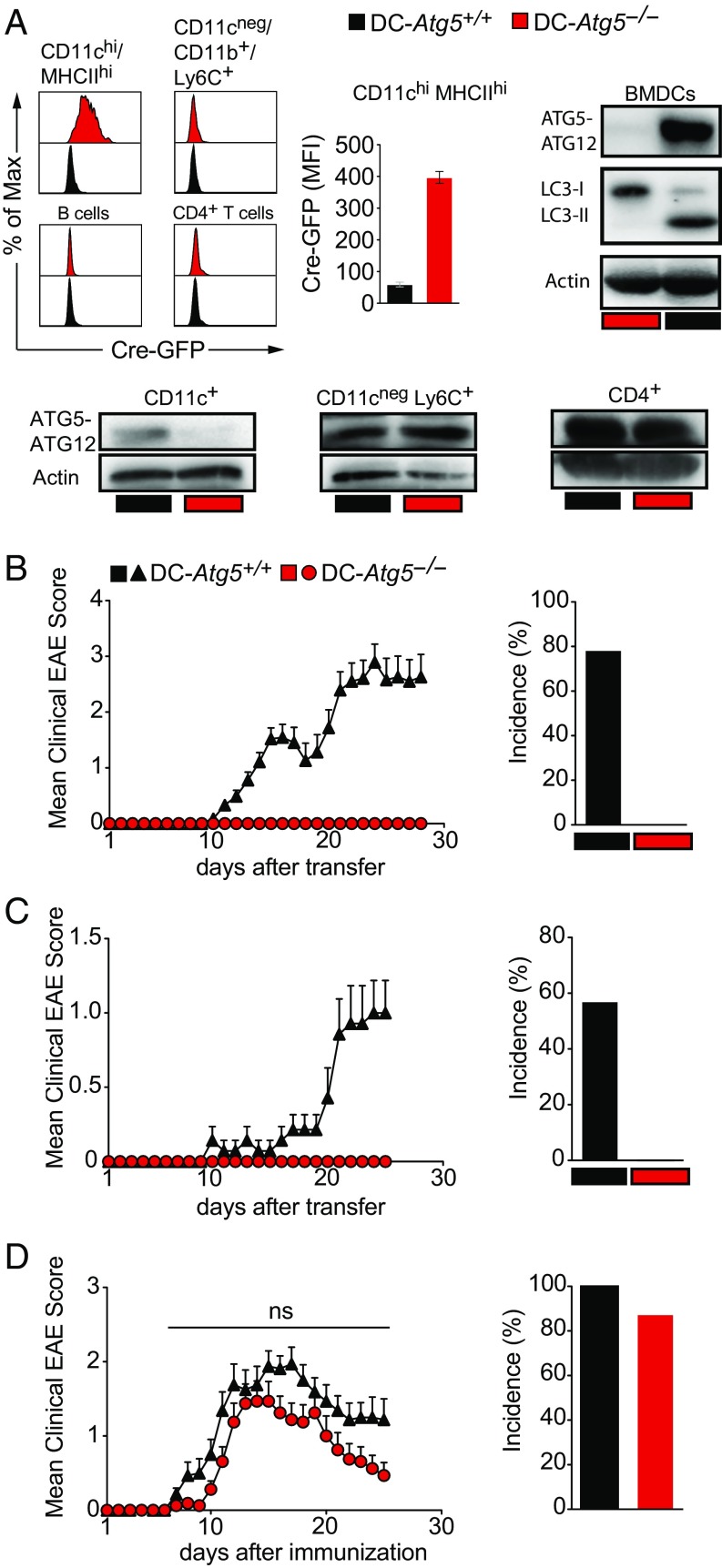
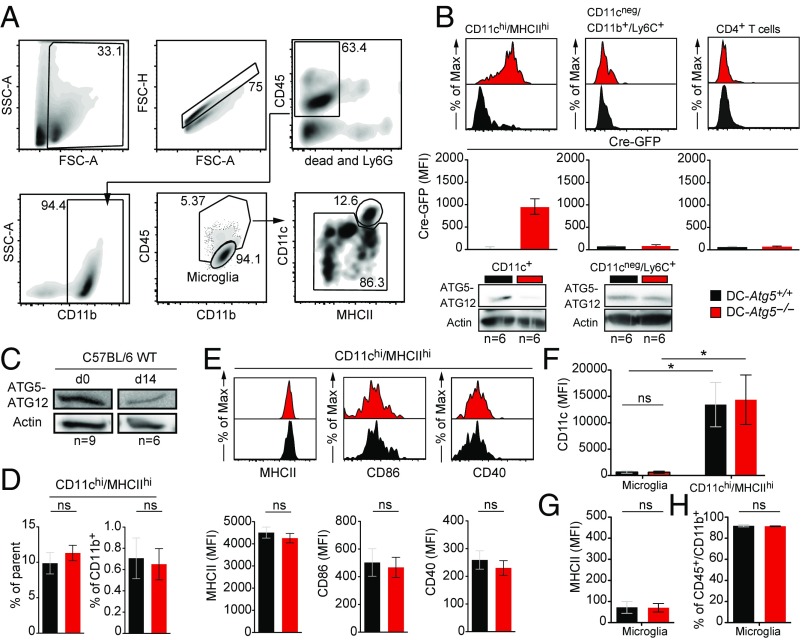
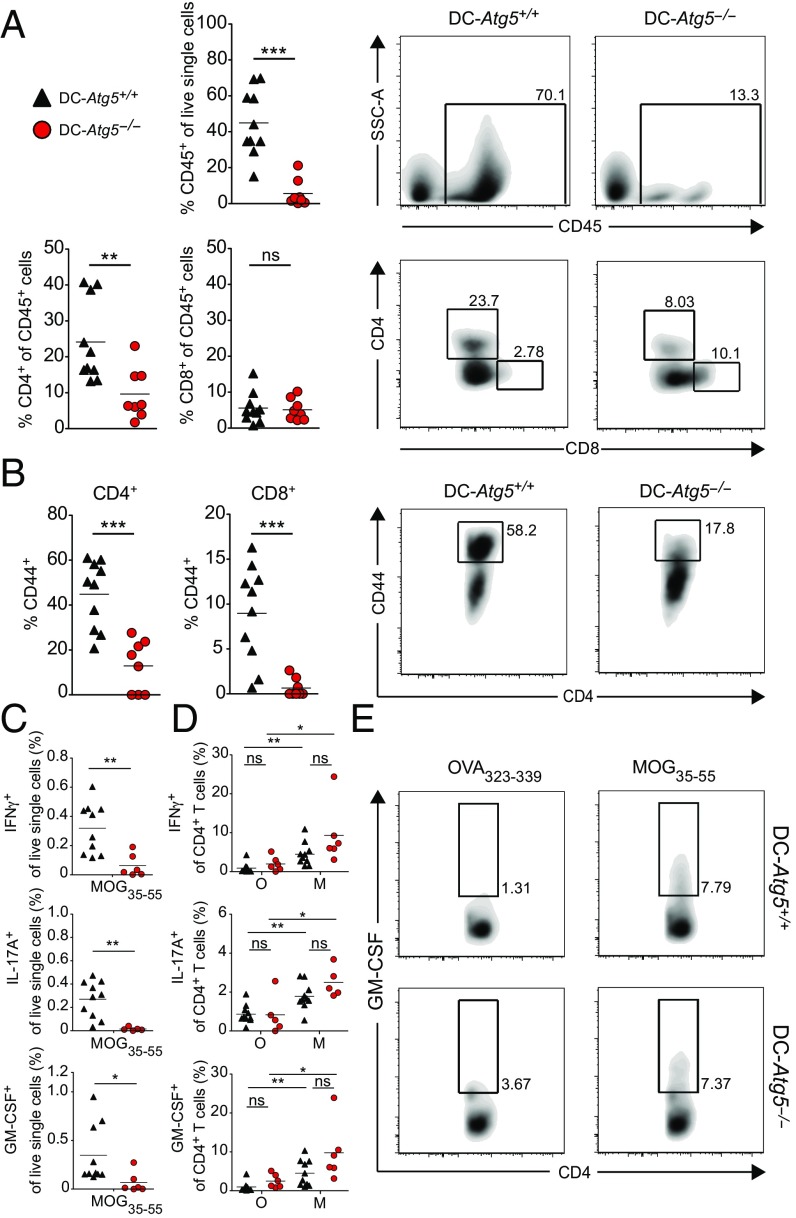
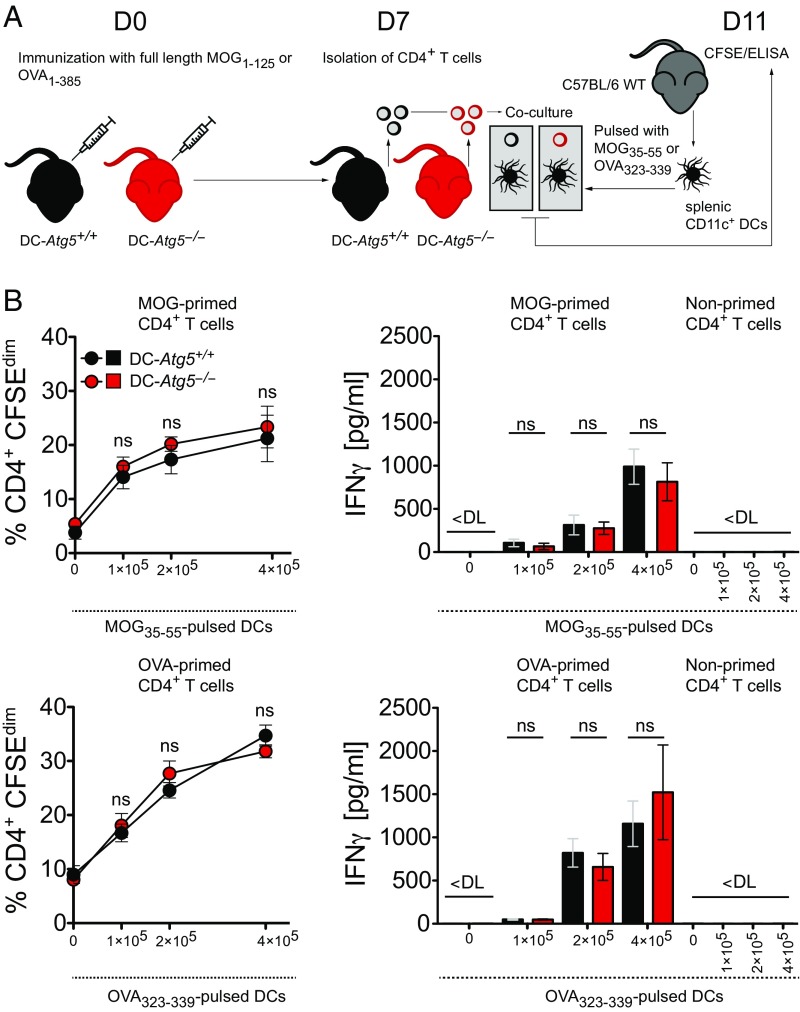
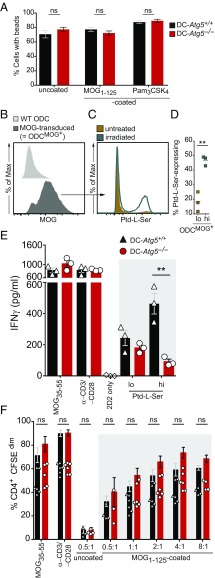
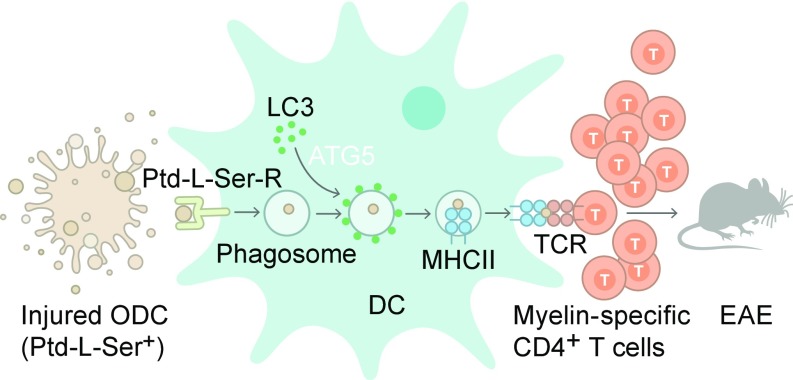
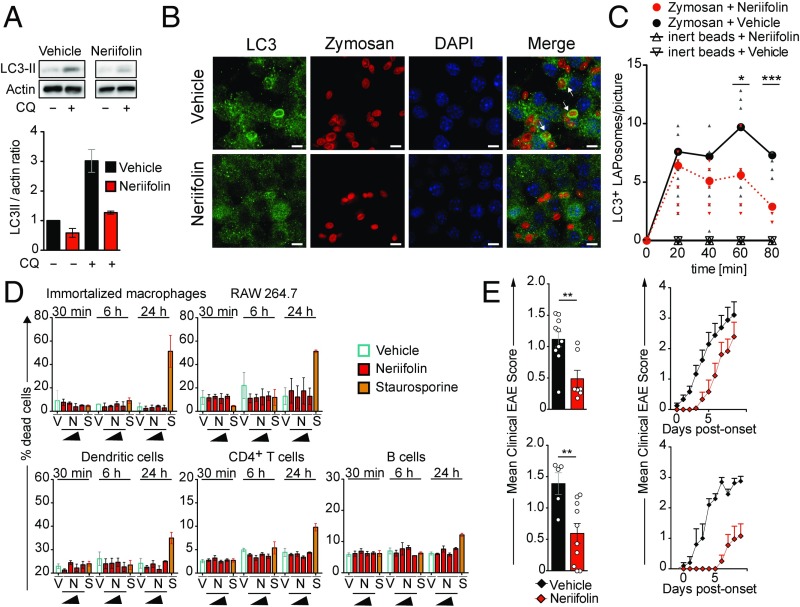
Although reactivation and accumulation of autoreactive CD4+ T cells within the CNS are considered to play a key role in the pathogenesis of multiple sclerosis (MS) and its animal model, experimental autoimmune encephalomyelitis (EAE), the mechanisms of how these cells recognize their target organ and induce sustained inflammation are incompletely understood. Here, we report that mice with conditional deletion of the essential autophagy protein ATG5 in classical dendritic cells (DCs), which are present at low frequencies in the nondiseased CNS, are completely resistant to EAE development following adoptive transfer of myelin-specific T cells and show substantially reduced in situ CD4+ T cell accumulation during the effector phase of the disease. Endogenous myelin peptide presentation to CD4+ T cells following phagocytosis of injured, phosphatidylserine-exposing oligodendroglial cells is abrogated in the absence of ATG5. Pharmacological inhibition of ATG-dependent phagocytosis by the cardiac glycoside neriifolin, an inhibitor of the Na+, K+-ATPase, delays the onset and reduces the clinical severity of EAE induced by myelin-specific CD4+ T cells. These findings link phagocytosis of injured oligodendrocytes, a pathological hallmark of MS lesions and during EAE, with myelin antigen processing and T cell pathogenicity, and identify ATG-dependent phagocytosis in DCs as a key regulator in driving autoimmune CD4+ T cell-mediated CNS damage.
Keywords: EAE; autophagy; multiple sclerosis; neuroinflammation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Beecham AH, et al. International Multiple Sclerosis Genetics Consortium (IMSGC); Wellcome Trust Case Control Consortium 2 (WTCCC2); International IBD Genetics Consortium (IIBDGC) Analysis of immune-related loci identifies 48 new susceptibility variants for multiple sclerosis. Nat Genet. 2013;45:1353–1360. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
