

Infrabarrels Are Layer 6 Circuit Modules in the Barrel Cortex that Link Long-Range Inputs and Outputs
- PMID: 29241536
- PMCID: PMC5736017
- DOI: 10.1016/j.celrep.2017.11.049
Infrabarrels Are Layer 6 Circuit Modules in the Barrel Cortex that Link Long-Range Inputs and Outputs
Abstract
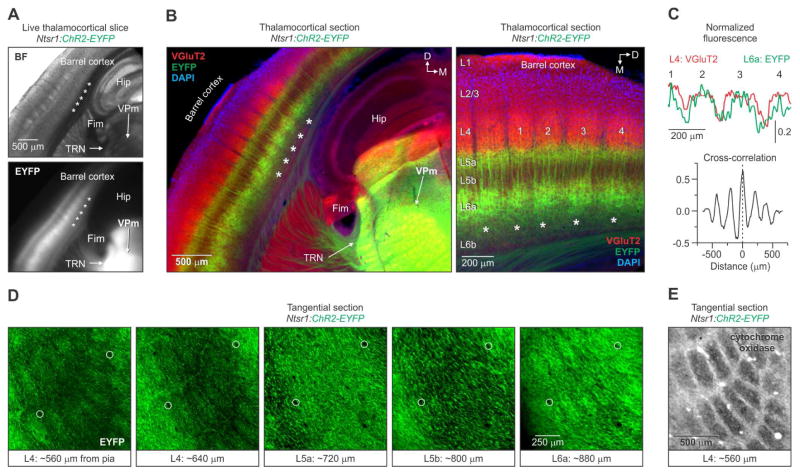
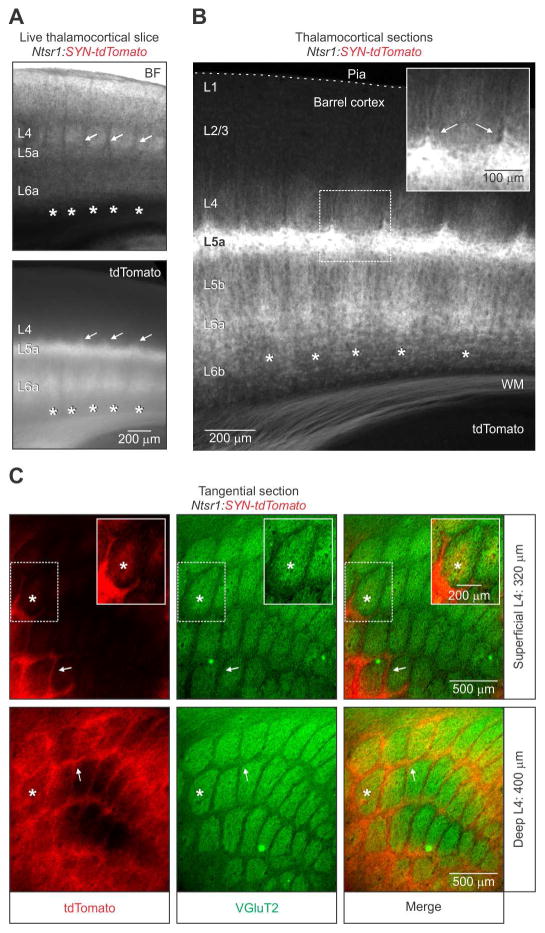
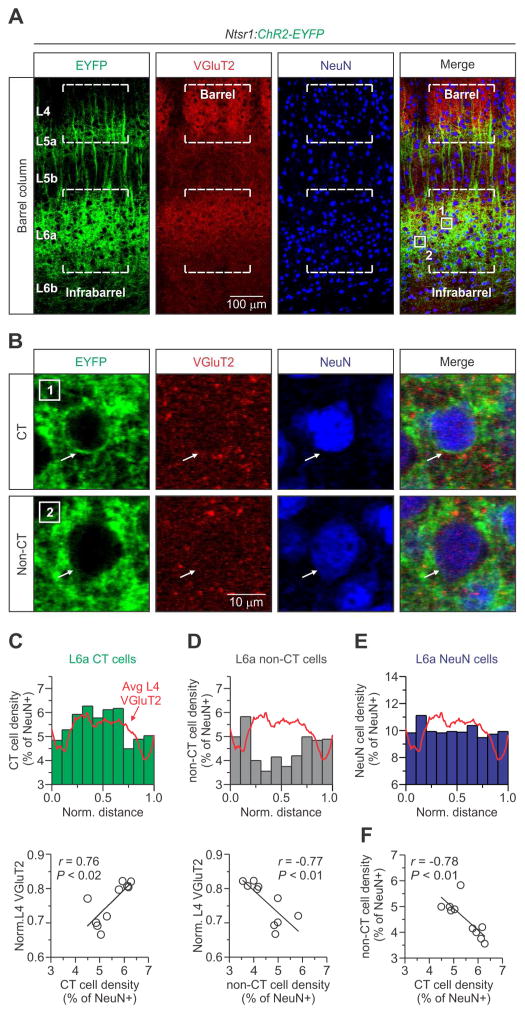
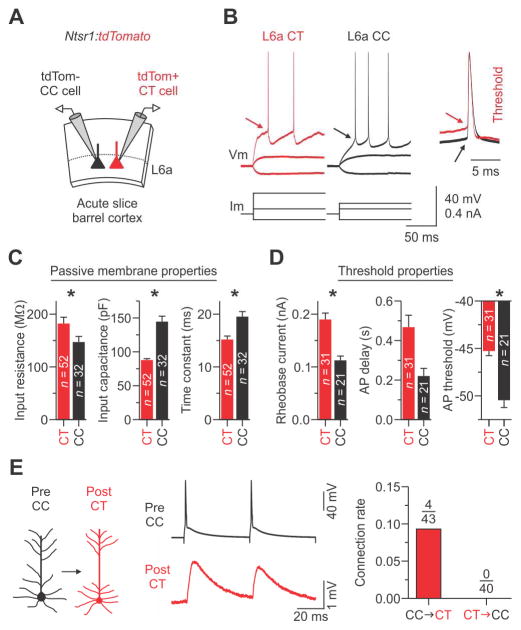
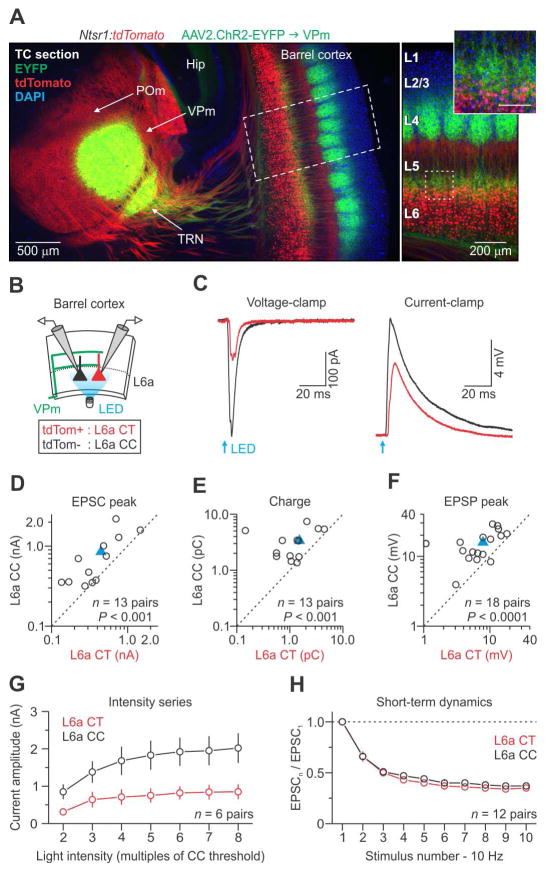
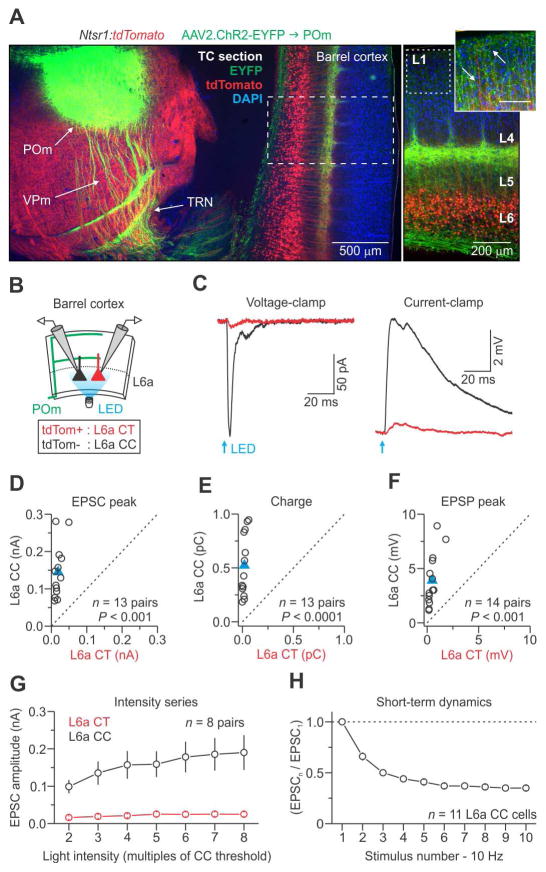
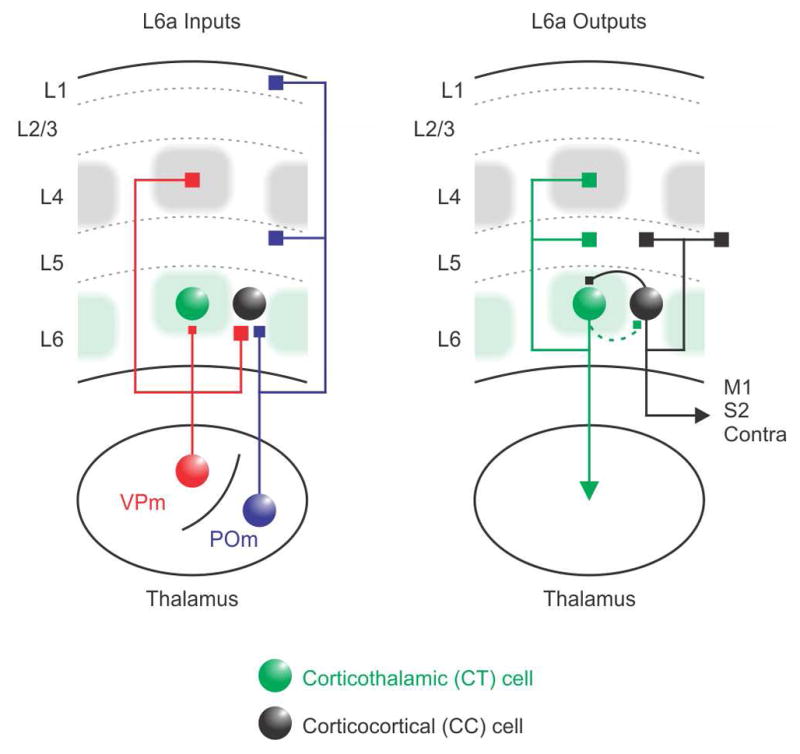
The rodent somatosensory cortex includes well-defined examples of cortical columns-the barrel columns-that extend throughout the cortical depth and are defined by discrete clusters of neurons in layer 4 (L4) called barrels. Using the cell-type-specific Ntsr1-Cre mouse line, we found that L6 contains infrabarrels, readily identifiable units that align with the L4 barrels. Corticothalamic (CT) neurons and their local axons cluster within the infrabarrels, whereas corticocortical (CC) neurons are densest between infrabarrels. Optogenetic experiments showed that CC cells received robust input from somatosensory thalamic nuclei, whereas CT cells received much weaker thalamic inputs. We also found that CT neurons are intrinsically less excitable, revealing that both synaptic and intrinsic mechanisms contribute to the low firing rates of CT neurons often reported in vivo. In summary, infrabarrels are discrete cortical circuit modules containing two partially separated excitatory networks that link long-distance thalamic inputs with specific outputs.
Keywords: channelrhodopsin; corticofugal; infragranular; layer 6; neocortex; septa; synapse; thalamocortical; thalamus; whisker.
Copyright © 2017 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Agmon A, Connors BW. Thalamocortical responses of mouse somatosensory (barrel) cortex in vitro. Neuroscience. 1991;41:365–379. - PubMed
-
- Ahissar E, Sosnik R, Haidarliu S. Transformation from temporal to rate coding in a somatosensory thalamocortical pathway. Nature. 2000;406:302–306. - PubMed
-
- Alloway KD. Information processing streams in rodent barrel cortex: the differential functions of barrel and septal circuits. Cerebral cortex. 2008;18:979–989. - PubMed
-
- Alloway KD, Zhang M, Chakrabarti S. Septal columns in rodent barrel cortex: functional circuits for modulating whisking behavior. J Comp Neurol. 2004;480:299–309. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
