

Conditional knockout mice for the distal appendage protein CEP164 reveal its essential roles in airway multiciliated cell differentiation
- PMID: 29244804
- PMCID: PMC5747467
- DOI: 10.1371/journal.pgen.1007128
Conditional knockout mice for the distal appendage protein CEP164 reveal its essential roles in airway multiciliated cell differentiation
Abstract
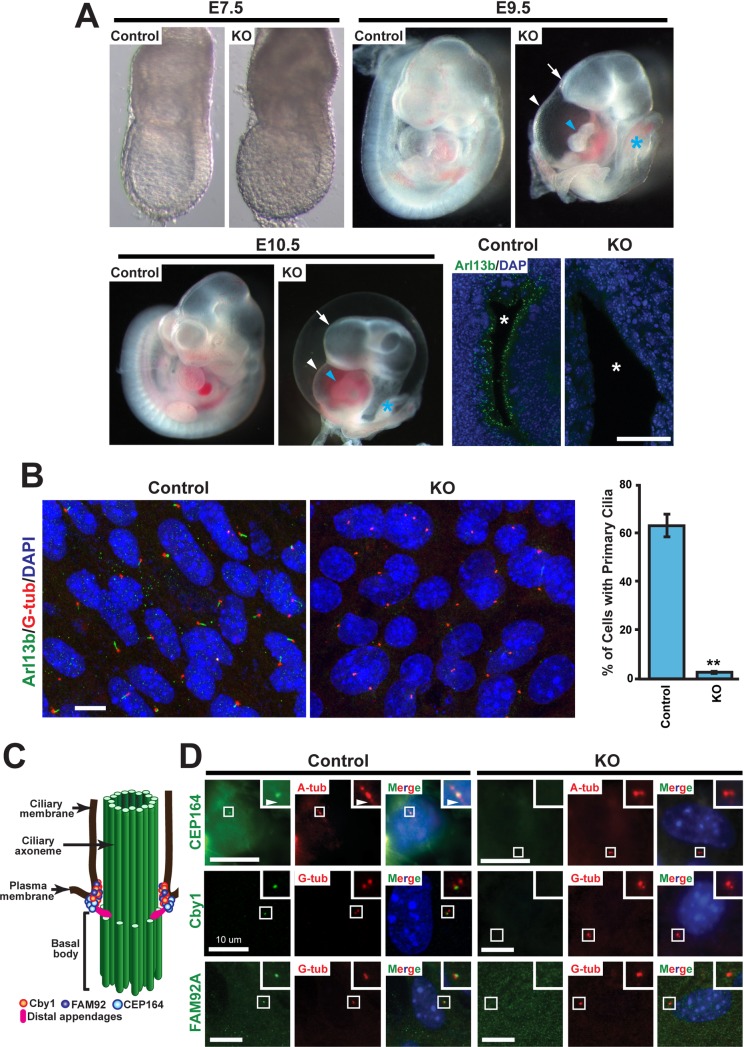
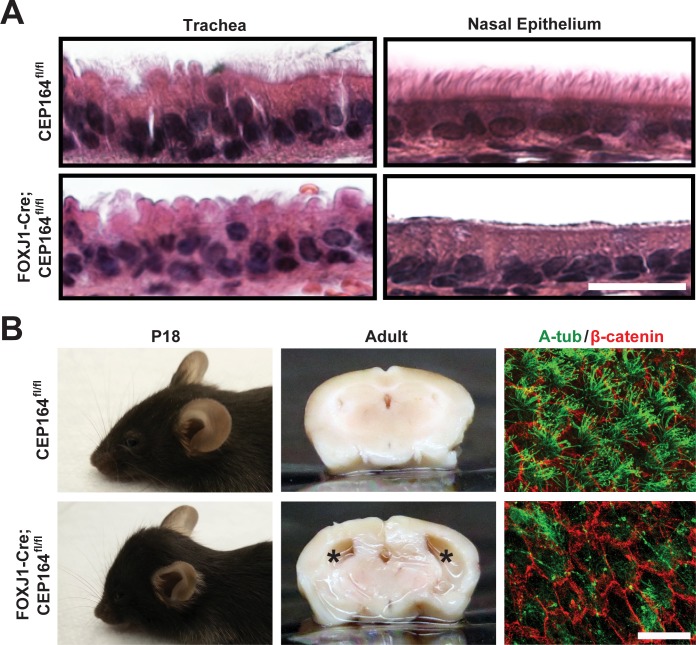
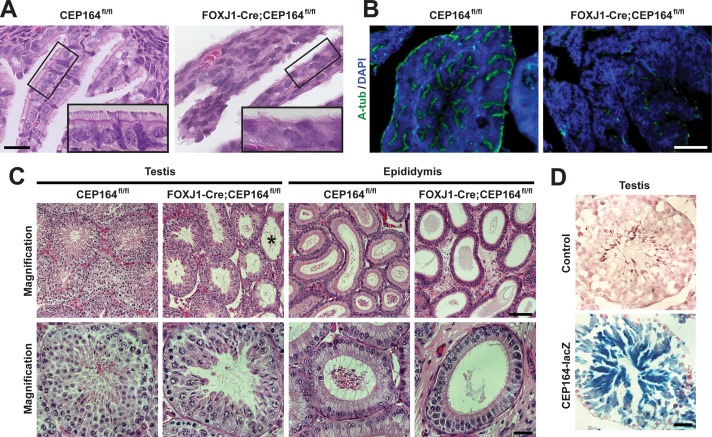
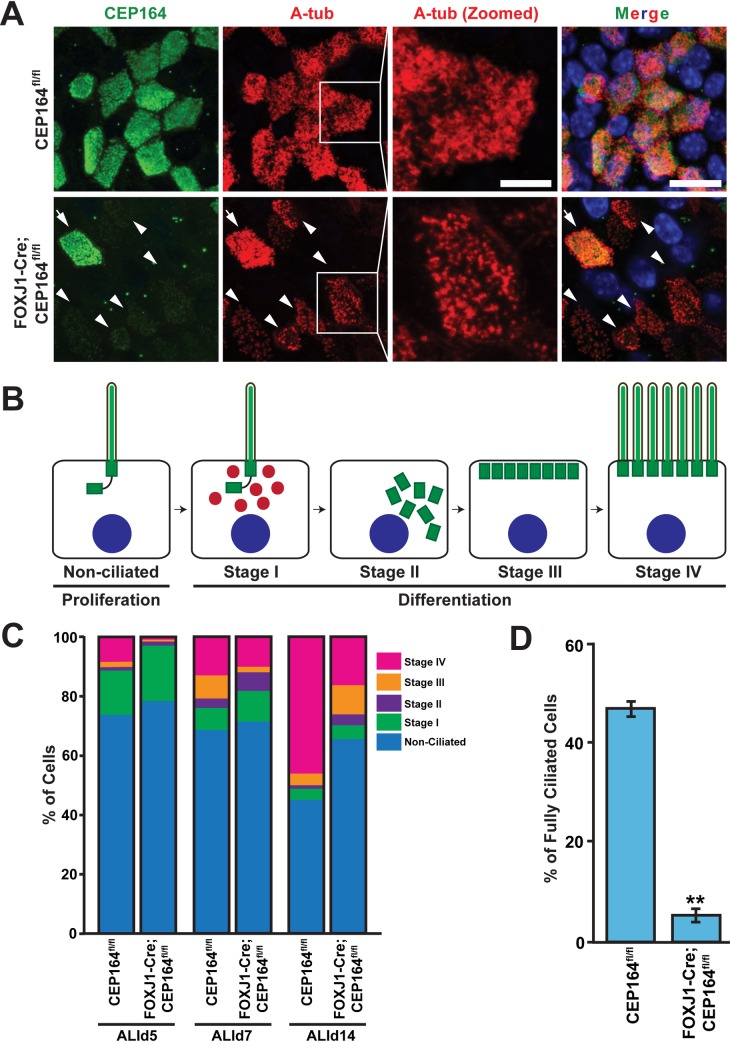
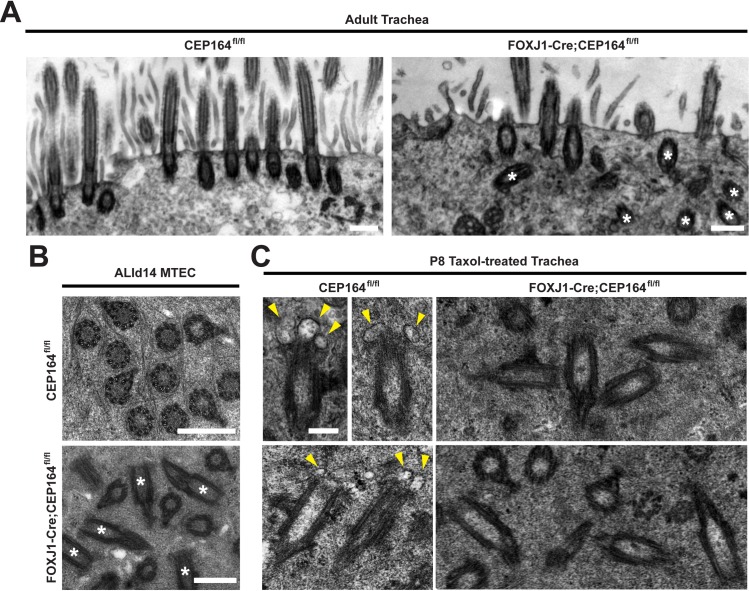
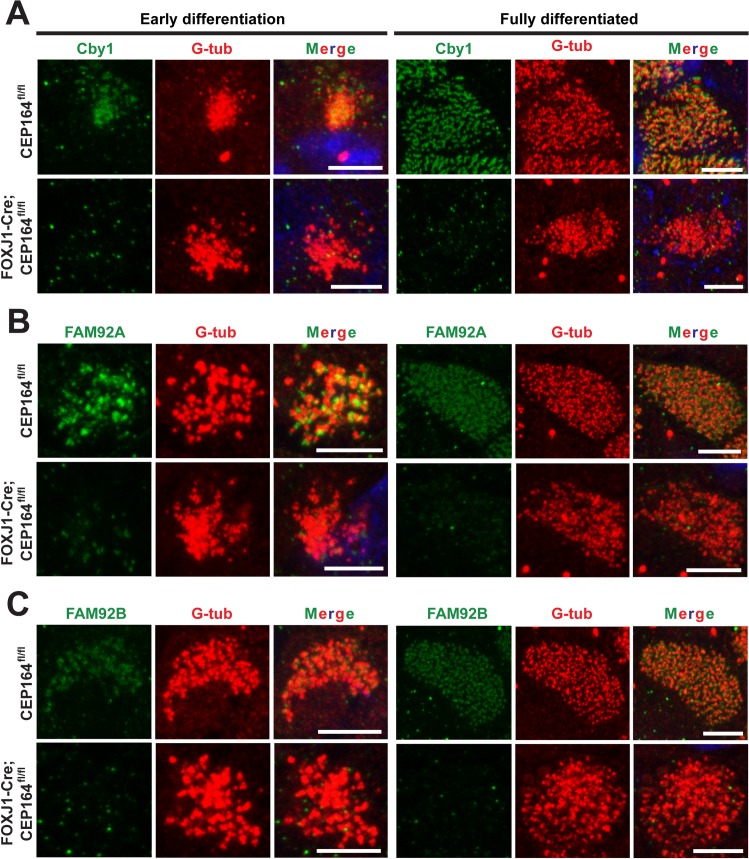
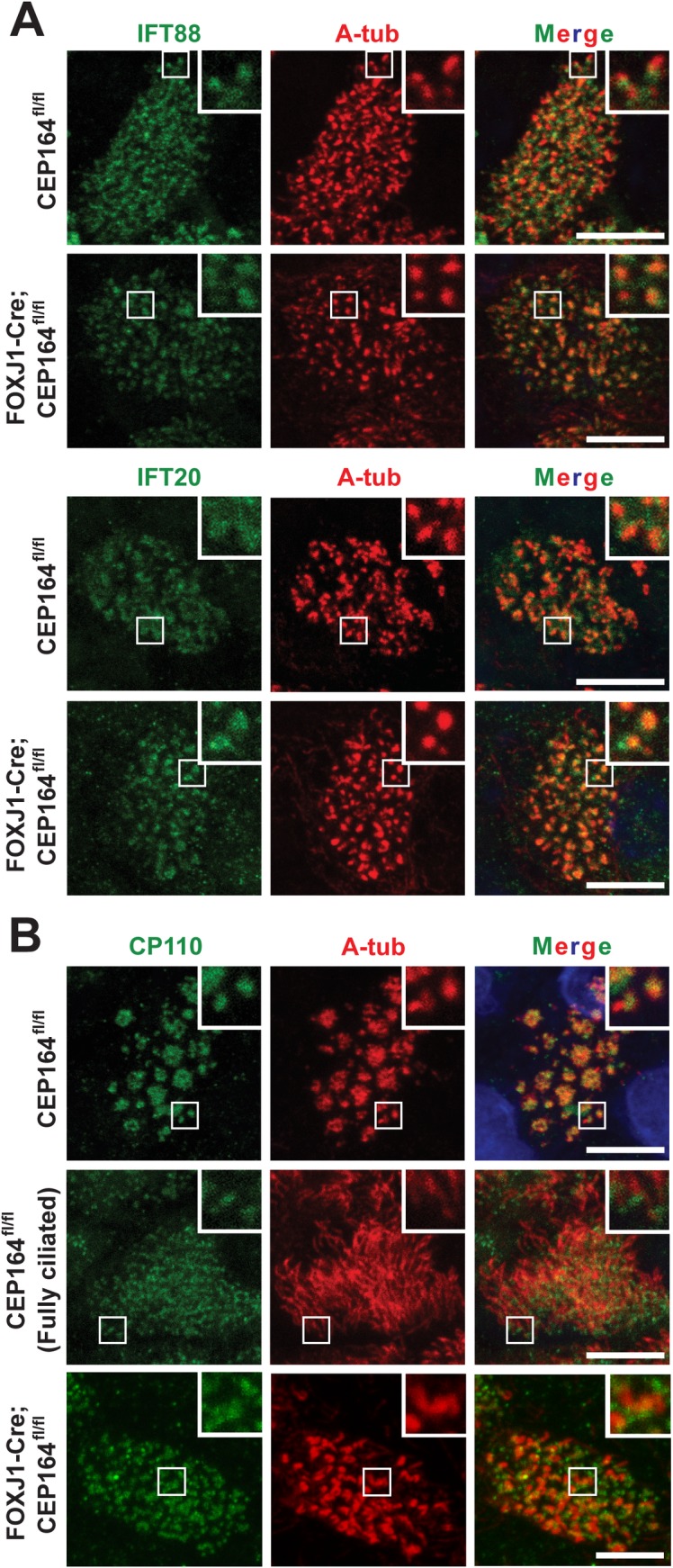
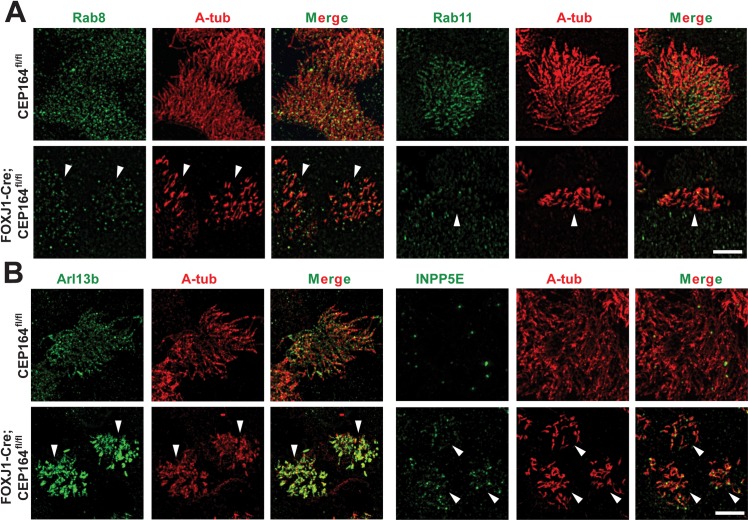
Multiciliated cells of the airways, brain ventricles, and female reproductive tract provide the motive force for mucociliary clearance, cerebrospinal fluid circulation, and ovum transport. Despite their clear importance to human biology and health, the molecular mechanisms underlying multiciliated cell differentiation are poorly understood. Prior studies implicate the distal appendage/transition fiber protein CEP164 as a central regulator of primary ciliogenesis; however, its role in multiciliogenesis remains unknown. In this study, we have generated a novel conditional mouse model that lacks CEP164 in multiciliated tissues and the testis. These mice show a profound loss of airway, ependymal, and oviduct multicilia and develop hydrocephalus and male infertility. Using primary cultures of tracheal multiciliated cells as a model system, we found that CEP164 is critical for multiciliogenesis, at least in part, via its regulation of small vesicle recruitment, ciliary vesicle formation, and basal body docking. In addition, CEP164 is necessary for the proper recruitment of another distal appendage/transition fiber protein Chibby1 (Cby1) and its binding partners FAM92A and FAM92B to the ciliary base in multiciliated cells. In contrast to primary ciliogenesis, CEP164 is dispensable for the recruitment of intraflagellar transport (IFT) components to multicilia. Finally, we provide evidence that CEP164 differentially controls the ciliary targeting of membrane-associated proteins, including the small GTPases Rab8, Rab11, and Arl13b, in multiciliated cells. Altogether, our studies unravel unique requirements for CEP164 in primary versus multiciliogenesis and suggest that CEP164 modulates the selective transport of membrane vesicles and their cargoes into the ciliary compartment in multiciliated cells. Furthermore, our mouse model provides a useful tool to gain physiological insight into diseases associated with defective multicilia.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
CEP164 is essential for efferent duct multiciliogenesis and male fertility.Reproduction. 2021 Jul 8;162(2):129-139. doi: 10.1530/REP-21-0042. Reproduction. 2021. PMID: 34085951 Free PMC article.
-
Chibby promotes ciliary vesicle formation and basal body docking during airway cell differentiation.J Cell Biol. 2014 Oct 13;207(1):123-37. doi: 10.1083/jcb.201406140. J Cell Biol. 2014. PMID: 25313408 Free PMC article.
-
BAR Domain-Containing FAM92 Proteins Interact with Chibby1 To Facilitate Ciliogenesis.Mol Cell Biol. 2016 Oct 13;36(21):2668-2680. doi: 10.1128/MCB.00160-16. Print 2016 Nov 1. Mol Cell Biol. 2016. PMID: 27528616 Free PMC article.
-
Essential Roles of Efferent Duct Multicilia in Male Fertility.Cells. 2022 Jan 20;11(3):341. doi: 10.3390/cells11030341. Cells. 2022. PMID: 35159149 Free PMC article. Review.
-
Role of DZIP1-CBY-FAM92 transition zone complex in the basal body to membrane attachment and ciliary budding.Biochem Soc Trans. 2020 Jun 30;48(3):1067-1075. doi: 10.1042/BST20191007. Biochem Soc Trans. 2020. PMID: 32491167 Review.
Cited by
-
CEP164 is essential for efferent duct multiciliogenesis and male fertility.Reproduction. 2021 Jul 8;162(2):129-139. doi: 10.1530/REP-21-0042. Reproduction. 2021. PMID: 34085951 Free PMC article.
-
CEP97 phosphorylation by Dyrk1a is critical for centriole separation during multiciliogenesis.J Cell Biol. 2022 Jan 3;221(1):e202102110. doi: 10.1083/jcb.202102110. Epub 2021 Nov 17. J Cell Biol. 2022. PMID: 34787650 Free PMC article.
-
Developmental and tissue-specific roles of mammalian centrosomes.FEBS J. 2025 Feb;292(4):709-726. doi: 10.1111/febs.17212. Epub 2024 Jun 27. FEBS J. 2025. PMID: 38935637 Free PMC article. Review.
-
Primary Cilia Influence Progenitor Function during Cortical Development.Cells. 2022 Sep 16;11(18):2895. doi: 10.3390/cells11182895. Cells. 2022. PMID: 36139475 Free PMC article. Review.
-
Gradual centriole maturation associates with the mitotic surveillance pathway in mouse development.EMBO Rep. 2021 Feb 3;22(2):e51127. doi: 10.15252/embr.202051127. Epub 2021 Jan 7. EMBO Rep. 2021. PMID: 33410253 Free PMC article.
References
-
- Goetz SC, Anderson KV. The primary cilium: a signalling centre during vertebrate development. Nature reviews Genetics. 2010;11(5):331–44. doi: 10.1038/nrg2774 ; PubMed Central PMCID: PMC3121168. - DOI - PMC - PubMed
-
- Nigg EA, Raff JW. Centrioles, centrosomes, and cilia in health and disease. Cell. 2009;139(4):663–78. doi: 10.1016/j.cell.2009.10.036 . - DOI - PubMed
-
- Hildebrandt F, Benzing T, Katsanis N. Ciliopathies. The New England journal of medicine. 2011;364(16):1533–43. doi: 10.1056/NEJMra1010172 ; PubMed Central PMCID: PMC3640822. - DOI - PMC - PubMed
-
- Brooks ER, Wallingford JB. Multiciliated Cells. Current biology: CB. 2014;24(19):R973–R82. doi: 10.1016/j.cub.2014.08.047 . - DOI - PMC - PubMed
-
- Spassky N, Meunier A. The development and functions of multiciliated epithelia. Nature reviews Molecular cell biology. 2017. doi: 10.1038/nrm.2017.21 . - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01HL107493 /NH/NIH HHS/United States
- F30HL129657 /NH/NIH HHS/United States
- T32GM008444-23 /NH/NIH HHS/United States
- R01 HL121791/HL/NHLBI NIH HHS/United States
- T32 GM008444/GM/NIGMS NIH HHS/United States
- R01 AI111605/AI/NIAID NIH HHS/United States
- R01AI111605 /NH/NIH HHS/United States
- S10 OD020096/OD/NIH HHS/United States
- RF1 AG057705/AG/NIA NIH HHS/United States
- R01 HL107493/HL/NHLBI NIH HHS/United States
- R01 AI130591/AI/NIAID NIH HHS/United States
- UH3 HL123429/HL/NHLBI NIH HHS/United States
- S10 OD016405/OD/NIH HHS/United States
- F30 HL129657/HL/NHLBI NIH HHS/United States
- R01HL121791 /NH/NIH HHS/United States
- UH3HL123429 /NH/NIH HHS/United States
- R56 HL107493/HL/NHLBI NIH HHS/United States
- R01AI130591 /NH/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
