

Optimal occlusion uniformly partitions red blood cells fluxes within a microvascular network
- PMID: 29244812
- PMCID: PMC5747476
- DOI: 10.1371/journal.pcbi.1005892
Optimal occlusion uniformly partitions red blood cells fluxes within a microvascular network
Abstract
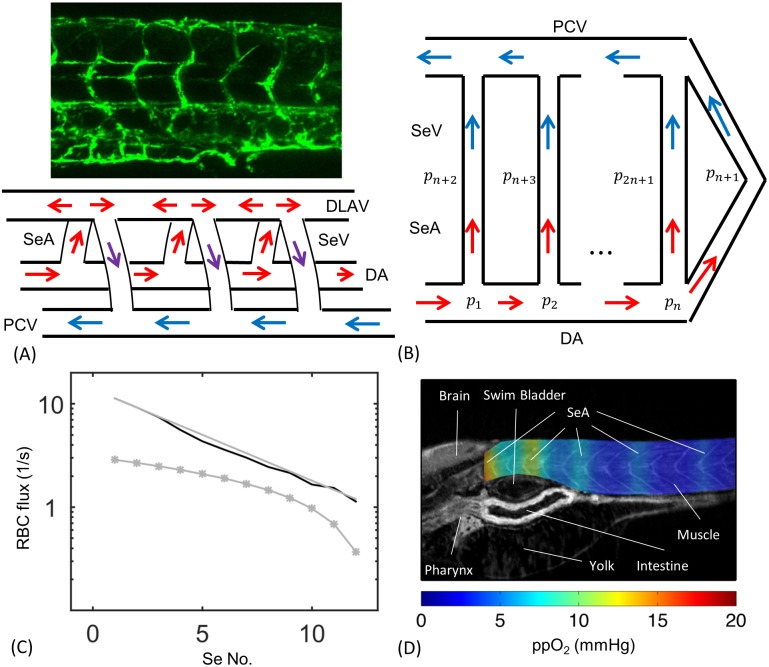
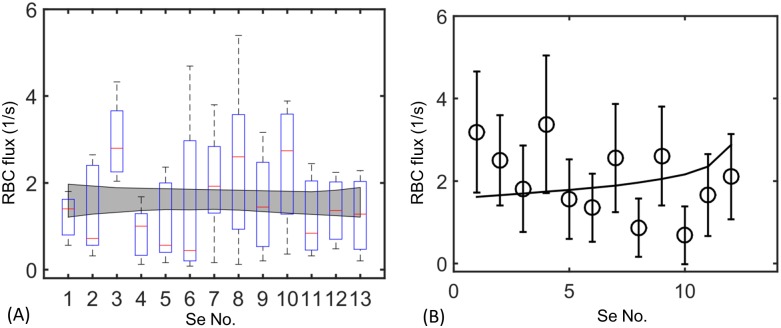
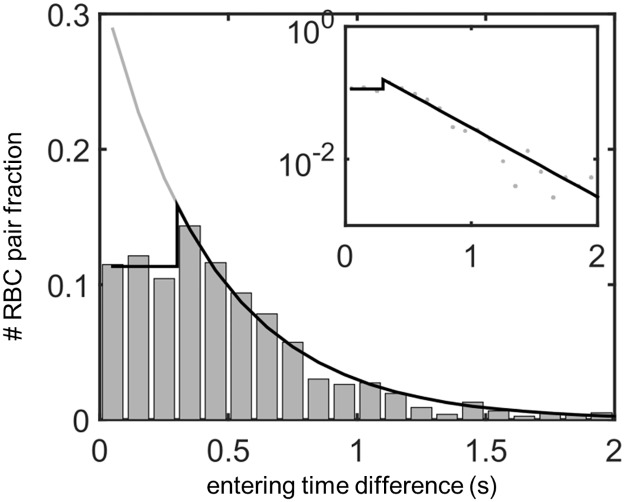
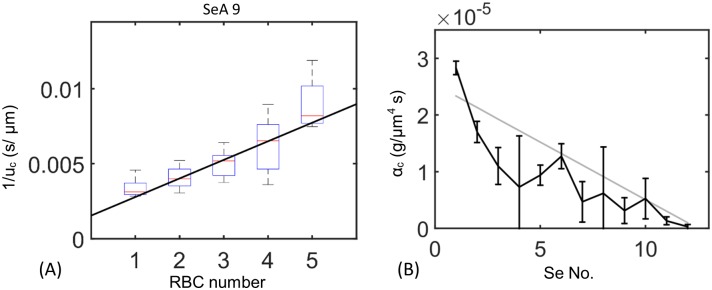
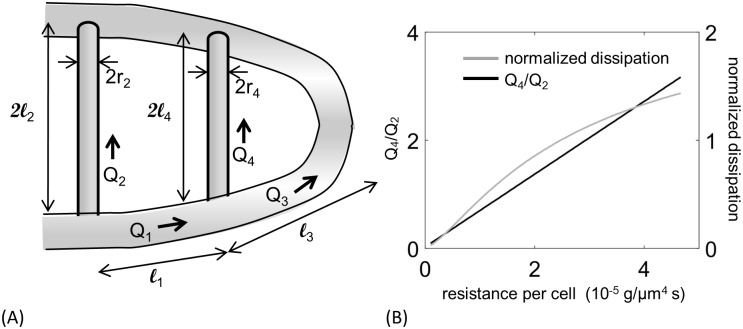
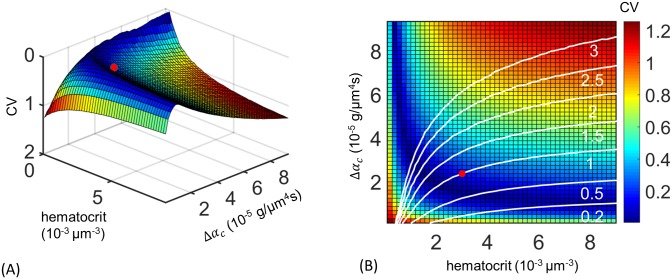
In animals, gas exchange between blood and tissues occurs in narrow vessels, whose diameter is comparable to that of a red blood cell. Red blood cells must deform to squeeze through these narrow vessels, transiently blocking or occluding the vessels they pass through. Although the dynamics of vessel occlusion have been studied extensively, it remains an open question why microvessels need to be so narrow. We study occlusive dynamics within a model microvascular network: the embryonic zebrafish trunk. We show that pressure feedbacks created when red blood cells enter the finest vessels of the trunk act together to uniformly partition red blood cells through the microvasculature. Using mathematical models as well as direct observation, we show that these occlusive feedbacks are tuned throughout the trunk network to prevent the vessels closest to the heart from short-circuiting the network. Thus occlusion is linked with another open question of microvascular function: how are red blood cells delivered at the same rate to each micro-vessel? Our analysis shows that tuning of occlusive feedbacks increase the total dissipation within the network by a factor of 11, showing that uniformity of flows rather than minimization of transport costs may be prioritized by the microvascular network.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Tomaiuolo G. Biomechanical properties of red blood cells in health and disease towards microfluidics. Biomicrofluidics. 2014;8(5):051501 doi: 10.1063/1.4895755 - DOI - PMC - PubMed
-
- West GB, Brown JH, Enquist BJ. A general model for the origin of allometric scaling laws in biology. Science. 1997;276(5309):122–126. doi: 10.1126/science.276.5309.122 - DOI - PubMed
-
- Savin T, Bandi M, Mahadevan L. Pressure-driven occlusive flow of a confined red blood cell. Soft matter. 2016;12(2):562–573. doi: 10.1039/C5SM01282A - DOI - PubMed
-
- Hall JE. Guyton and Hall textbook of medical physiology. Elsevier Health Sciences; 2015.
-
- Wan J, Ristenpart WD, Stone HA. Dynamics of shear-induced ATP release from red blood cells. Proceedings of the National Academy of Sciences. 2008;105(43):16432–16437. doi: 10.1073/pnas.0805779105 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
