

WNT Signaling in Cardiac and Vascular Disease
- PMID: 29247129
- PMCID: PMC6040091
- DOI: 10.1124/pr.117.013896
WNT Signaling in Cardiac and Vascular Disease
Abstract
WNT signaling is an elaborate and complex collection of signal transduction pathways mediated by multiple signaling molecules. WNT signaling is critically important for developmental processes, including cell proliferation, differentiation and tissue patterning. Little WNT signaling activity is present in the cardiovascular system of healthy adults, but reactivation of the pathway is observed in many pathologies of heart and blood vessels. The high prevalence of these pathologies and their significant contribution to human disease burden has raised interest in WNT signaling as a potential target for therapeutic intervention. In this review, we first will focus on the constituents of the pathway and their regulation and the different signaling routes. Subsequently, the role of WNT signaling in cardiovascular development is addressed, followed by a detailed discussion of its involvement in vascular and cardiac disease. After highlighting the crosstalk between WNT, transforming growth factor-β and angiotensin II signaling, and the emerging role of WNT signaling in the regulation of stem cells, we provide an overview of drugs targeting the pathway at different levels. From the combined studies we conclude that, despite the sometimes conflicting experimental data, a general picture is emerging that excessive stimulation of WNT signaling adversely affects cardiovascular pathology. The rapidly increasing collection of drugs interfering at different levels of WNT signaling will allow the evaluation of therapeutic interventions in the pathway in relevant animal models of cardiovascular diseases and eventually in patients in the near future, translating the outcomes of the many preclinical studies into a clinically relevant context.
Copyright © 2017 by The Author(s).
Figures
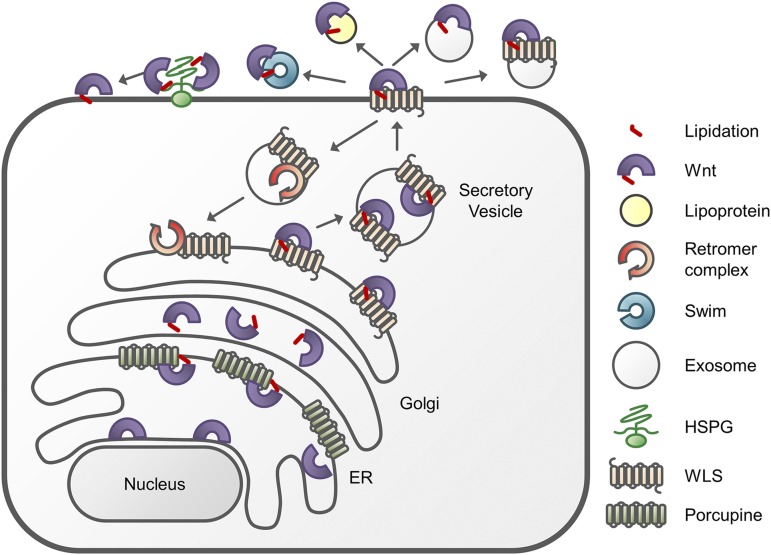
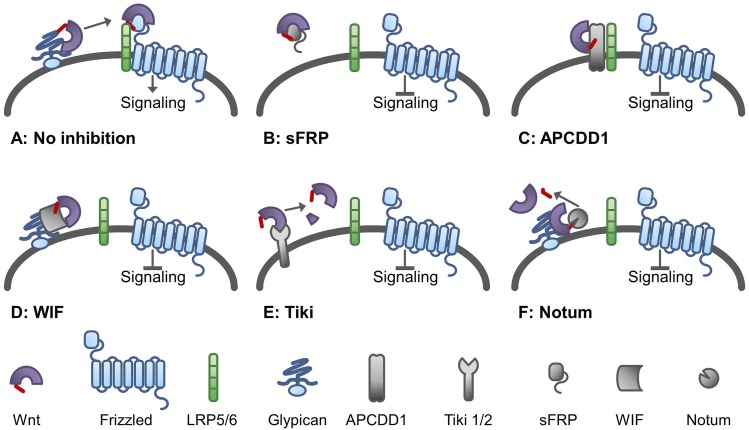
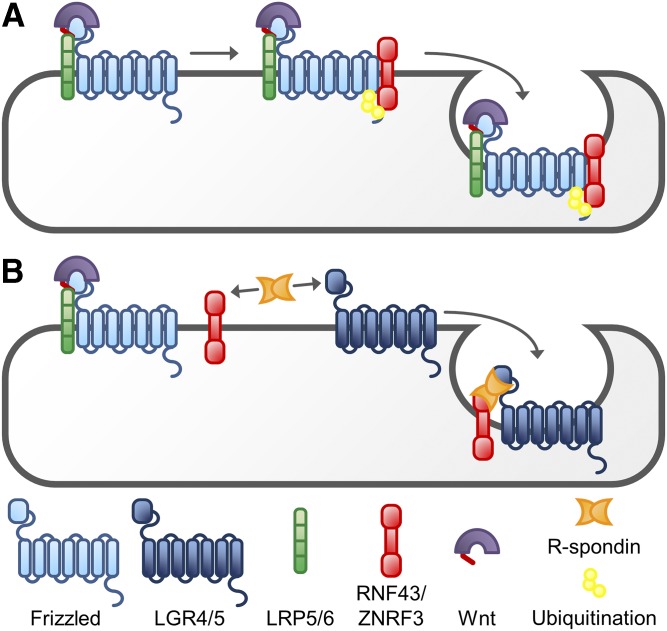
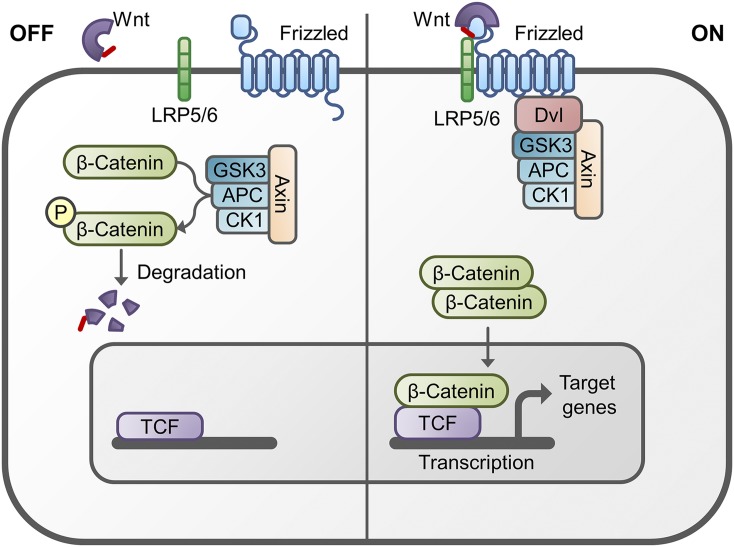
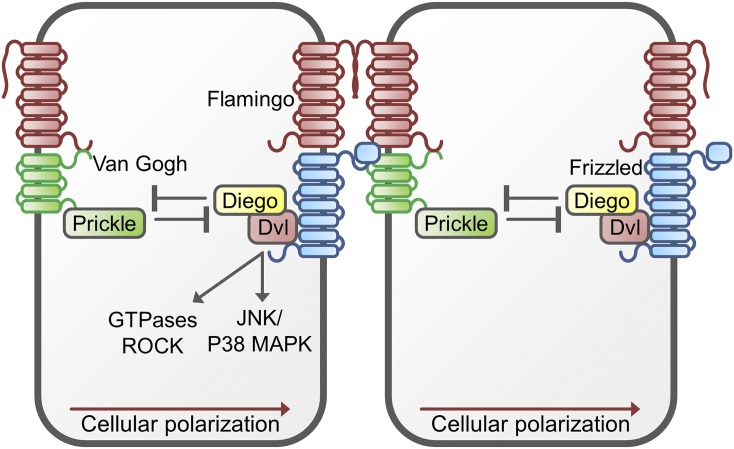
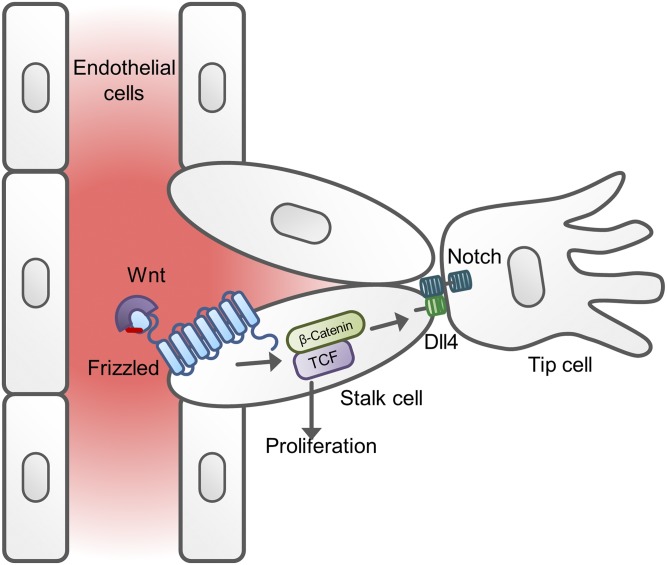
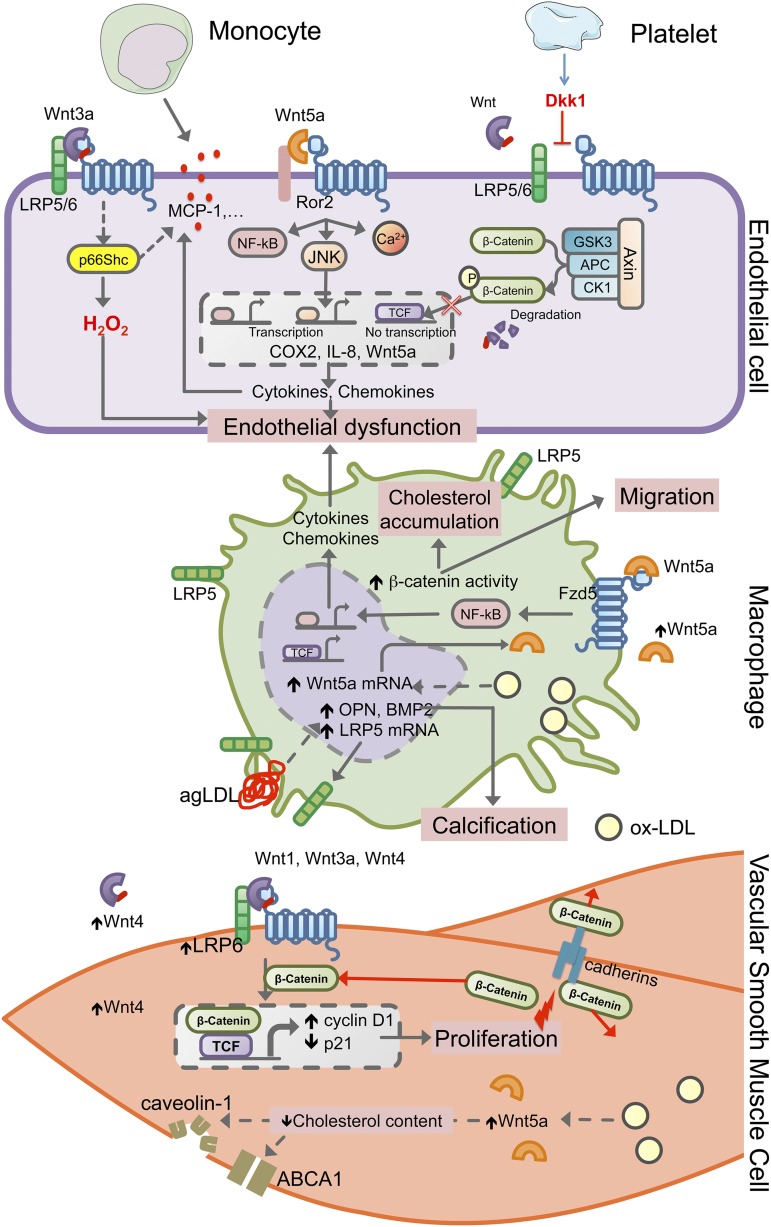
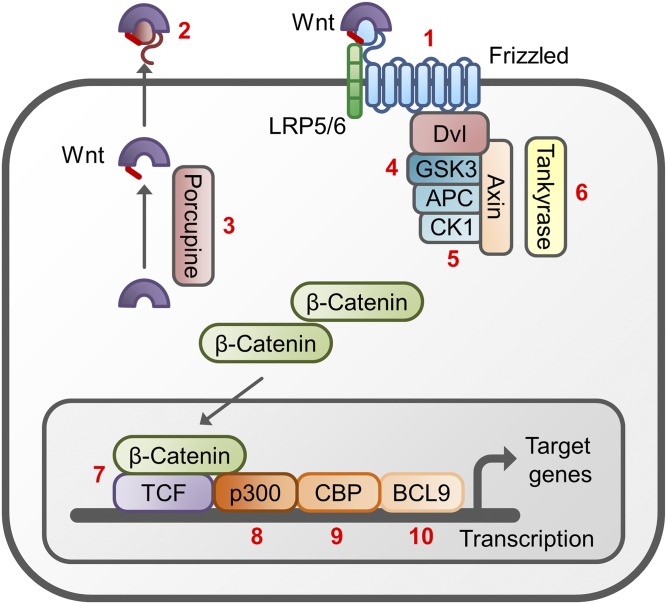
References
-
- Abbott NJ, Patabendige AA, Dolman DE, Yusof SR, Begley DJ. (2010) Structure and function of the blood-brain barrier. Neurobiol Dis 37:13–25. - PubMed
-
- Ahn J, Lee H, Kim S, Ha T. (2010) Curcumin-induced suppression of adipogenic differentiation is accompanied by activation of Wnt/beta-catenin signaling. Am J Physiol Cell Physiol 298:C1510–C1516. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
