

Structural dynamics of protein S1 on the 70S ribosome visualized by ensemble cryo-EM
- PMID: 29247757
- PMCID: PMC5866760
- DOI: 10.1016/j.ymeth.2017.12.004
Structural dynamics of protein S1 on the 70S ribosome visualized by ensemble cryo-EM
Abstract
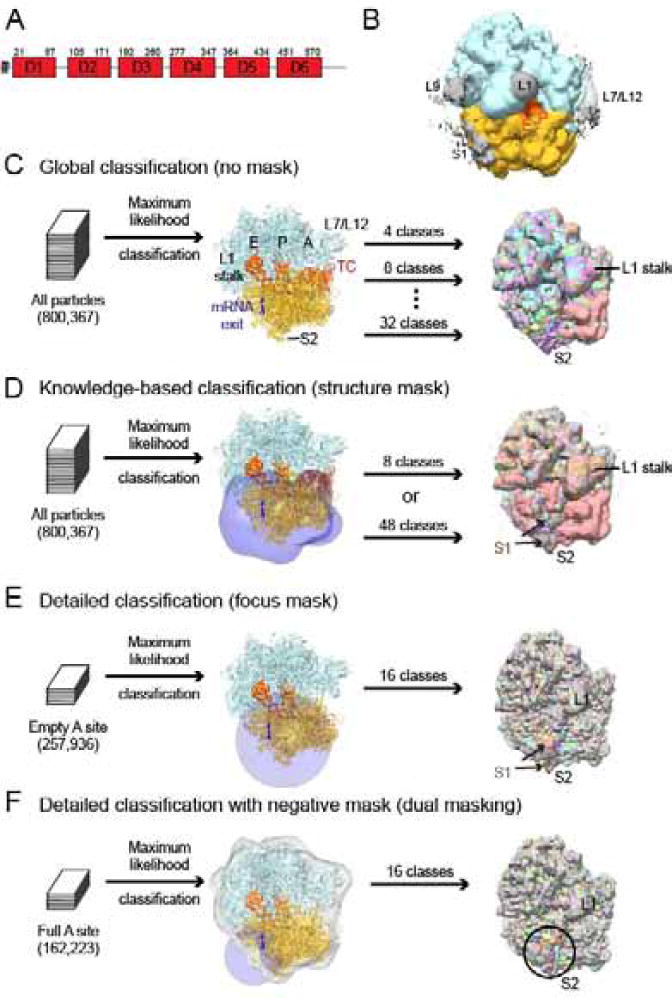
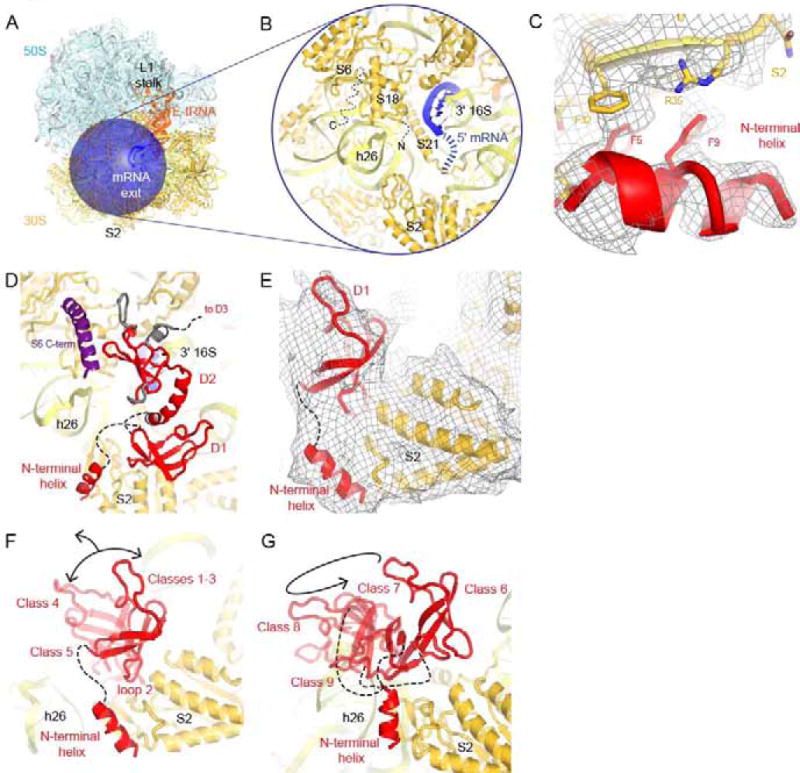
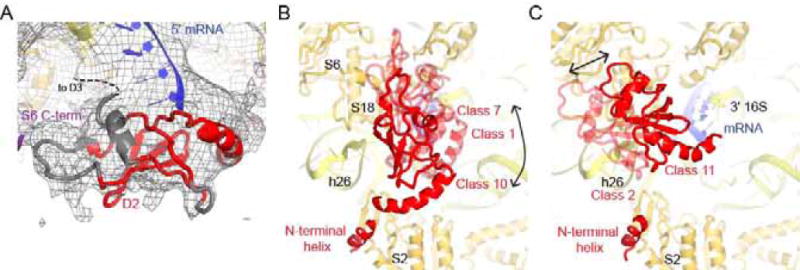
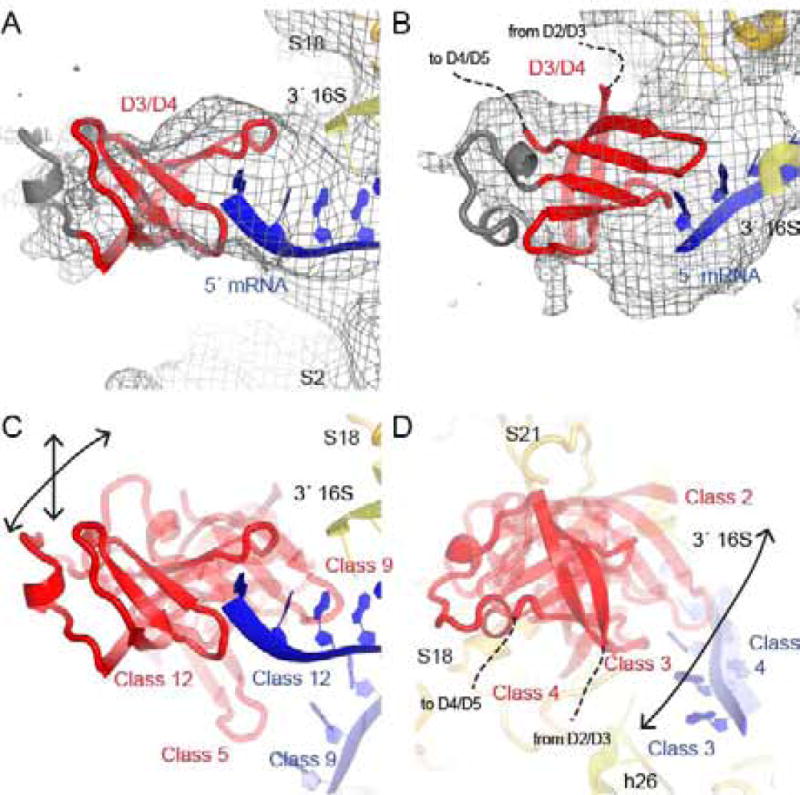
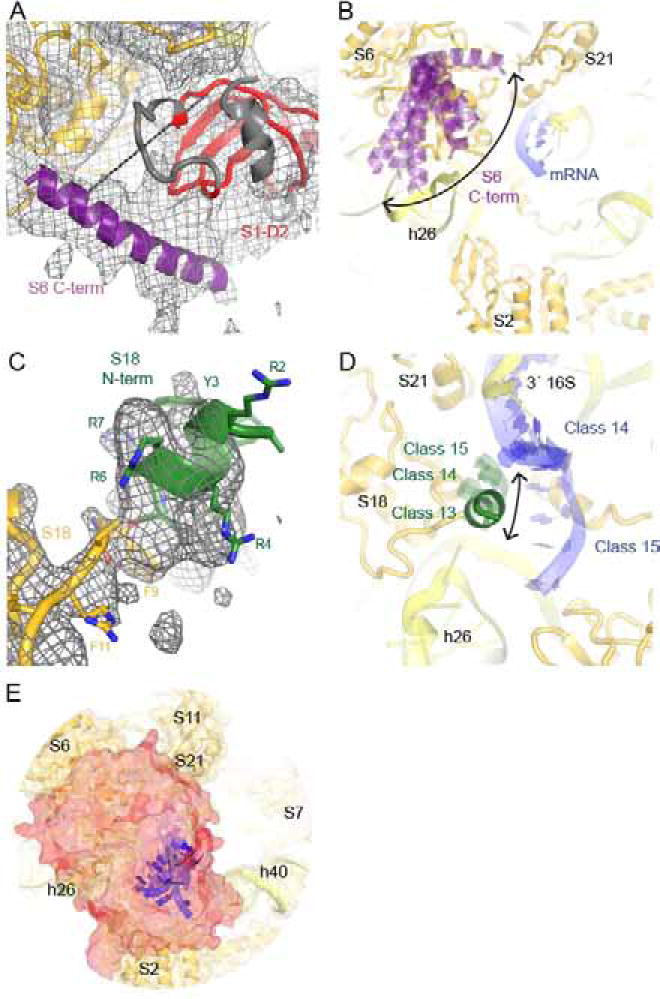
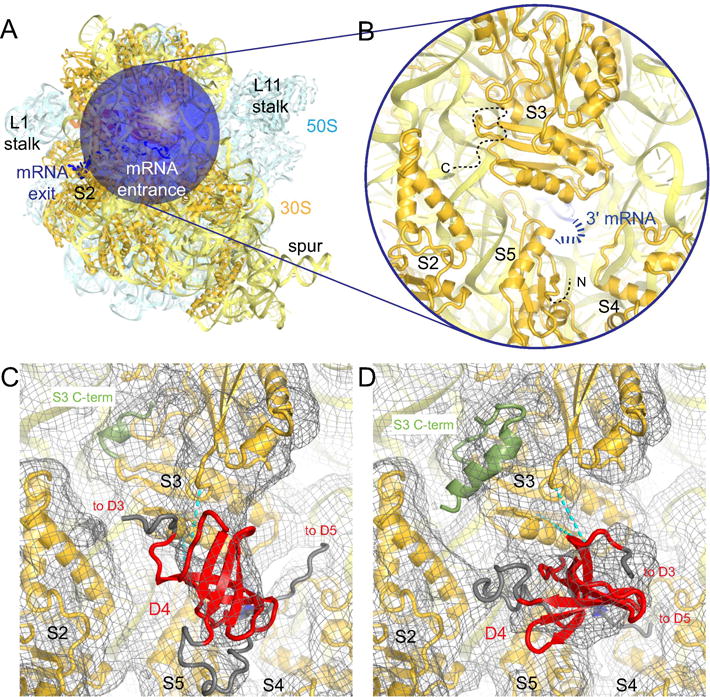
Bacterial ribosomal protein S1 is the largest and highly flexible protein of the 30S subunit, and one of a few core ribosomal proteins for which a complete structure is lacking. S1 is thought to participate in transcription and translation. Best understood is the role of S1 in facilitating translation of mRNAs with structured 5' UTRs. Here, we present cryo-EM analyses of the 70S ribosome that reveal multiple conformations of S1. Based on comparison of several 3D maximum likelihood classification approaches in Frealign, we propose a streamlined strategy for visualizing a highly dynamic component of a large macromolecular assembly that itself exhibits high compositional and conformational heterogeneity. The resulting maps show how S1 docks at the ribosomal protein S2 near the mRNA exit channel. The globular OB-fold domains sample a wide area around the mRNA exit channel and interact with mobile tails of proteins S6 and S18. S1 also interacts with the mRNA entrance channel, where an OB-fold domain can be localized near S3 and S5. Our analyses suggest that S1 cooperates with other ribosomal proteins to form a dynamic mesh near the mRNA exit and entrance channels to modulate the binding, folding and movement of mRNA.
Keywords: 3D maximum-likelihood classification; Electron cryo-microscopy; Ribosome; Rps1.
Copyright © 2017 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures
Similar articles
-
A method for differentiating proteins from nucleic acids in intermediate-resolution density maps: cryo-electron microscopy defines the quaternary structure of the Escherichia coli 70S ribosome.Structure. 2000 Sep 15;8(9):937-48. doi: 10.1016/s0969-2126(00)00185-4. Structure. 2000. PMID: 10986461
-
Structure of the bacterial ribosome at 2 Å resolution.Elife. 2020 Sep 14;9:e60482. doi: 10.7554/eLife.60482. Elife. 2020. PMID: 32924932 Free PMC article.
-
Conformational variability in Escherichia coli 70S ribosome as revealed by 3D cryo-electron microscopy.Int J Biochem Cell Biol. 1999 Jan;31(1):243-54. doi: 10.1016/s1357-2725(98)00149-6. Int J Biochem Cell Biol. 1999. PMID: 10216957
-
Cryo-electron microscopy as an investigative tool: the ribosome as an example.Bioessays. 2001 Aug;23(8):725-32. doi: 10.1002/bies.1102. Bioessays. 2001. PMID: 11494321 Review.
-
Ribosome dynamics: insights from atomic structure modeling into cryo-electron microscopy maps.Annu Rev Biophys Biomol Struct. 2006;35:299-317. doi: 10.1146/annurev.biophys.35.040405.101950. Annu Rev Biophys Biomol Struct. 2006. PMID: 16689638 Review.
Cited by
-
Unstructured 5'-tails act through ribosome standby to override inhibitory structure at ribosome binding sites.Nucleic Acids Res. 2018 May 4;46(8):4188-4199. doi: 10.1093/nar/gky073. Nucleic Acids Res. 2018. PMID: 29420821 Free PMC article.
-
Conformational switch in the ribosomal protein S1 guides unfolding of structured RNAs for translation initiation.Nucleic Acids Res. 2018 Nov 16;46(20):10917-10929. doi: 10.1093/nar/gky746. Nucleic Acids Res. 2018. PMID: 30124944 Free PMC article.
-
Investigation of the Relationship between the S1 Domain and Its Molecular Functions Derived from Studies of the Tertiary Structure.Molecules. 2019 Oct 13;24(20):3681. doi: 10.3390/molecules24203681. Molecules. 2019. PMID: 31614904 Free PMC article.
-
Multiple Antimicrobial Effects of Hybrid Peptides Synthesized Based on the Sequence of Ribosomal S1 Protein from Staphylococcus aureus.Int J Mol Sci. 2022 Jan 4;23(1):524. doi: 10.3390/ijms23010524. Int J Mol Sci. 2022. PMID: 35008951 Free PMC article.
-
NMR structure of the Vibrio vulnificus ribosomal protein S1 domains D3 and D4 provides insights into molecular recognition of single-stranded RNAs.Nucleic Acids Res. 2021 Jul 21;49(13):7753-7764. doi: 10.1093/nar/gkab562. Nucleic Acids Res. 2021. PMID: 34223902 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
