

R-2HG Exhibits Anti-tumor Activity by Targeting FTO/m6A/MYC/CEBPA Signaling
- PMID: 29249359
- PMCID: PMC5766423
- DOI: 10.1016/j.cell.2017.11.031
R-2HG Exhibits Anti-tumor Activity by Targeting FTO/m6A/MYC/CEBPA Signaling
Abstract
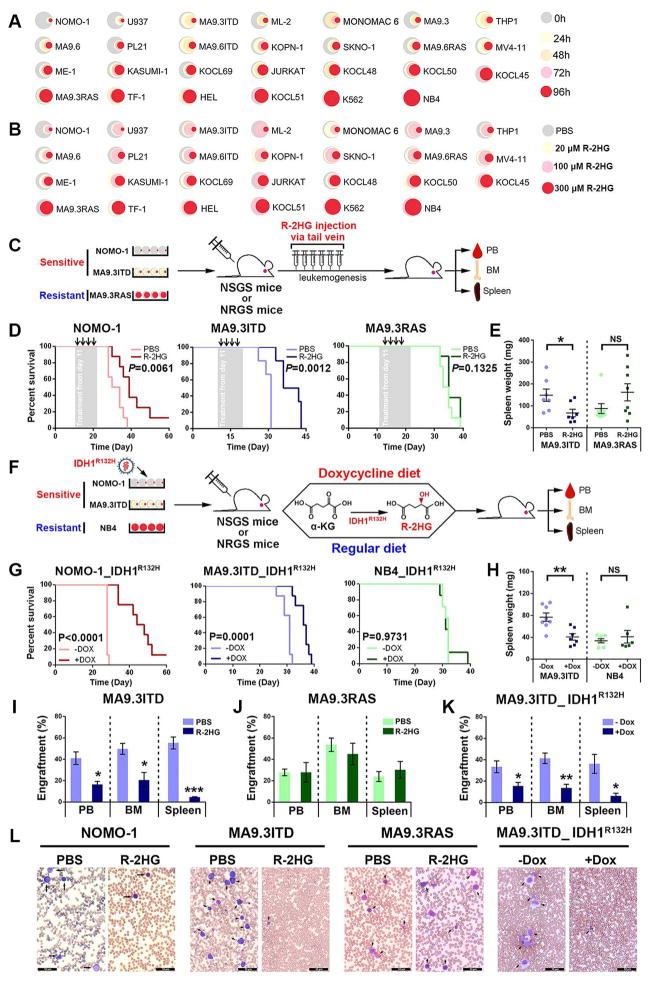
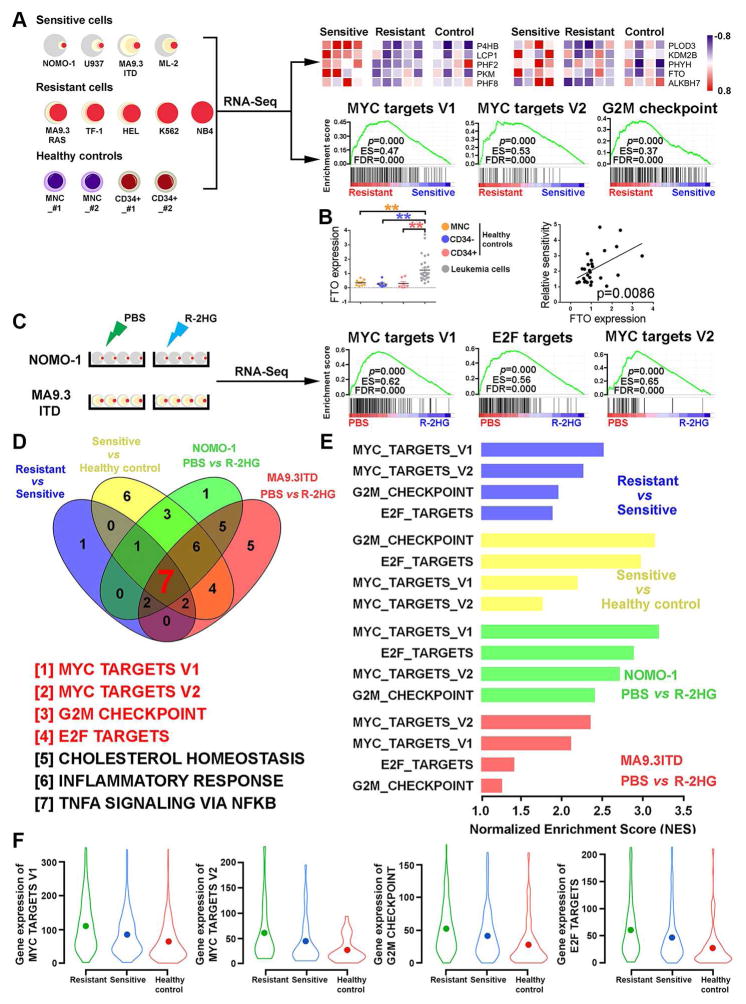
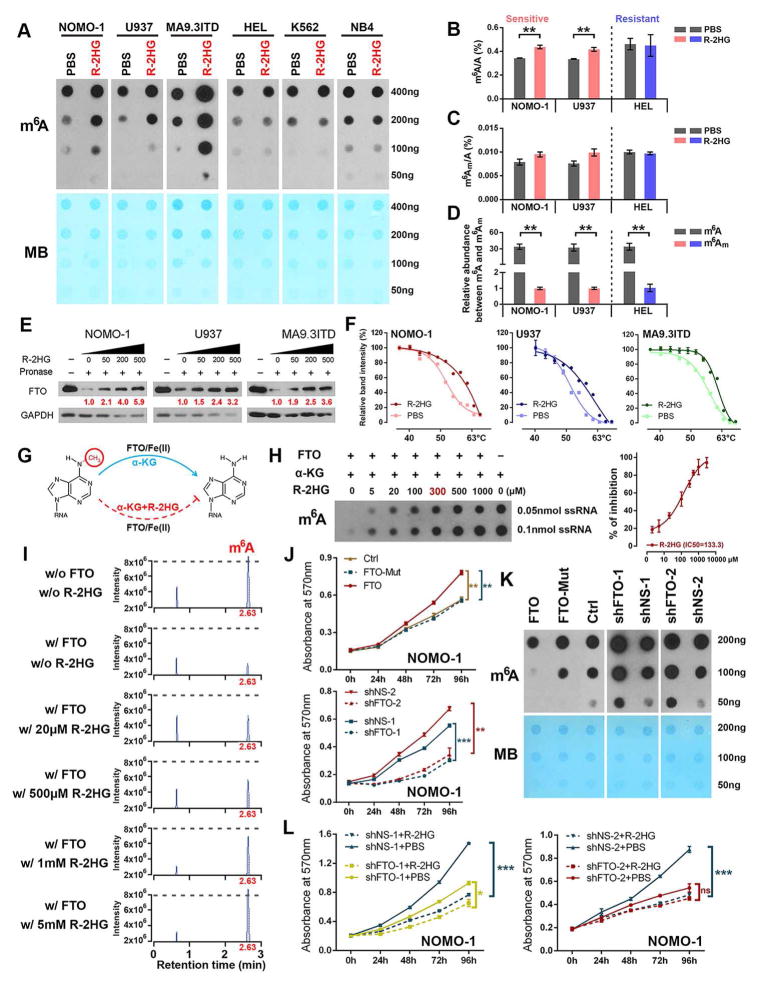
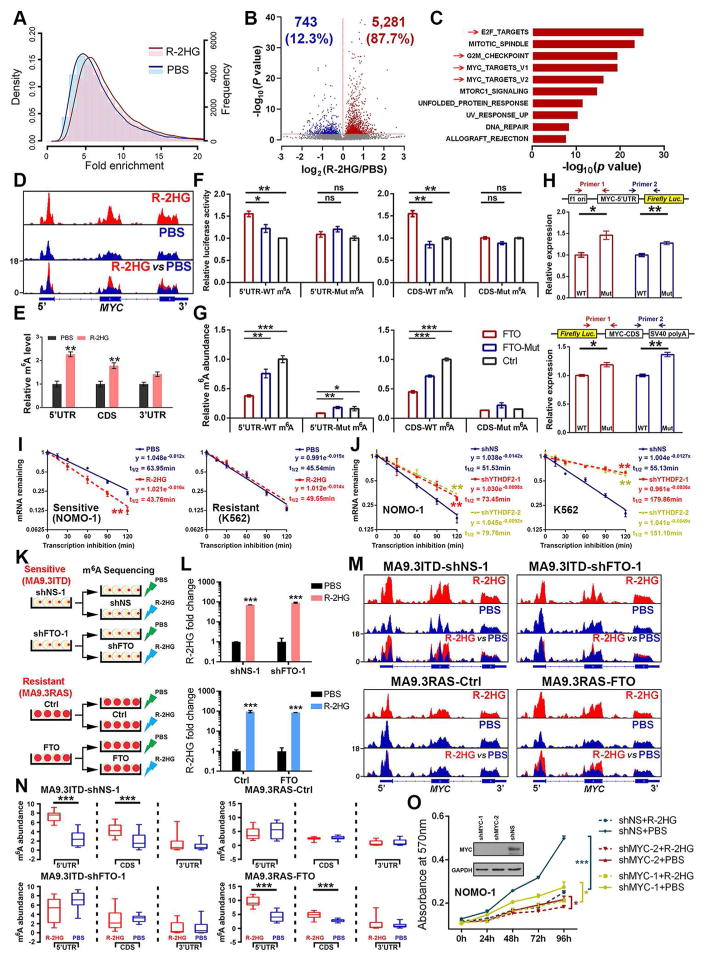
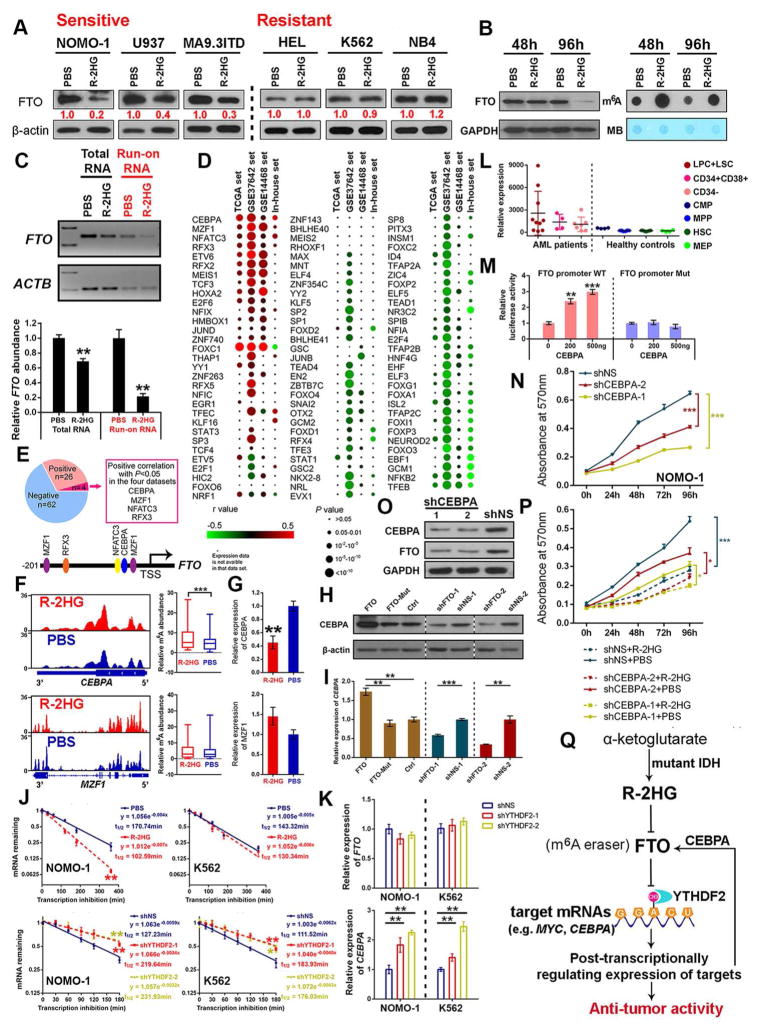
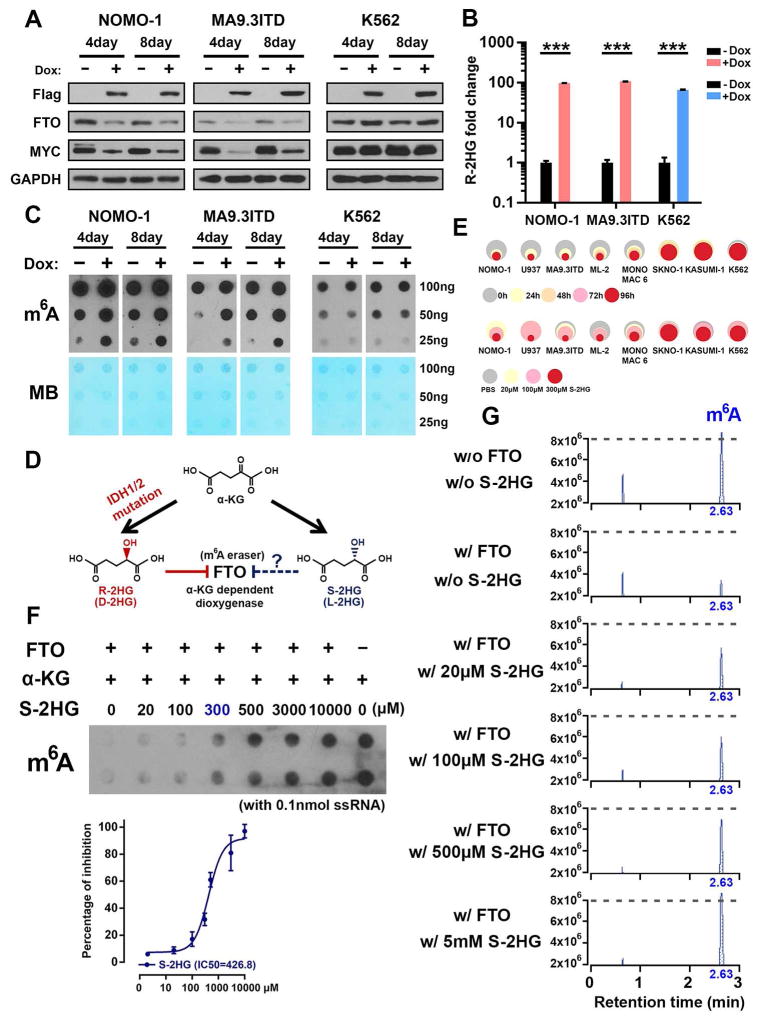
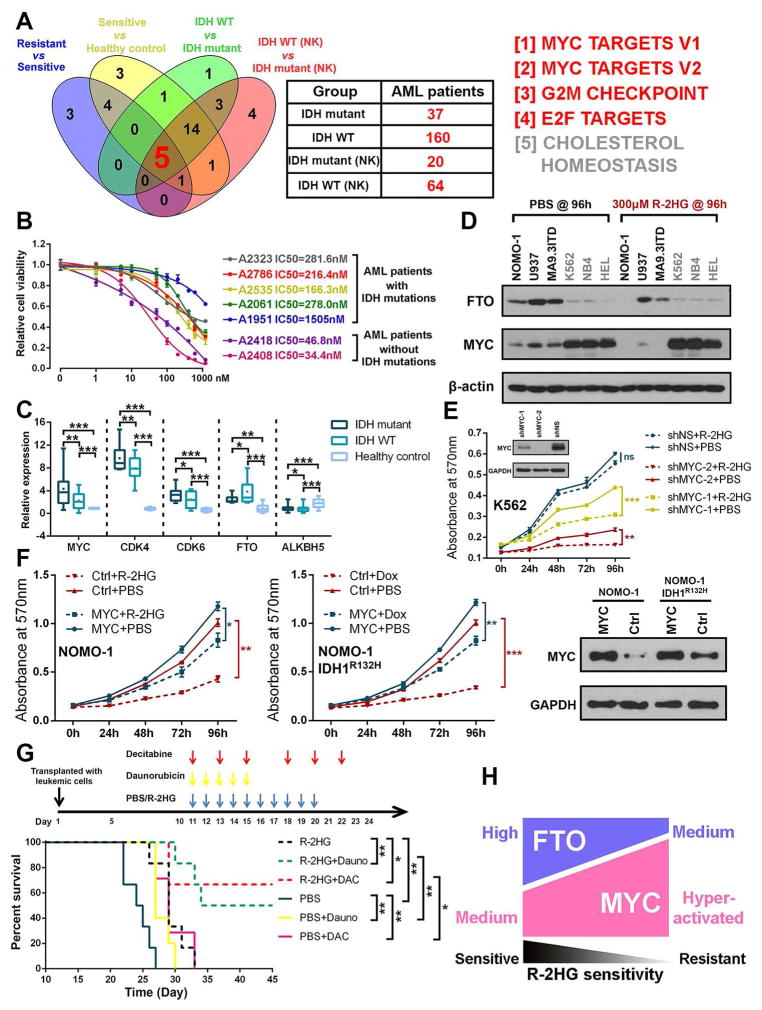
R-2-hydroxyglutarate (R-2HG), produced at high levels by mutant isocitrate dehydrogenase 1/2 (IDH1/2) enzymes, was reported as an oncometabolite. We show here that R-2HG also exerts a broad anti-leukemic activity in vitro and in vivo by inhibiting leukemia cell proliferation/viability and by promoting cell-cycle arrest and apoptosis. Mechanistically, R-2HG inhibits fat mass and obesity-associated protein (FTO) activity, thereby increasing global N6-methyladenosine (m6A) RNA modification in R-2HG-sensitive leukemia cells, which in turn decreases the stability of MYC/CEBPA transcripts, leading to the suppression of relevant pathways. Ectopically expressed mutant IDH1 and S-2HG recapitulate the effects of R-2HG. High levels of FTO sensitize leukemic cells to R-2HG, whereas hyperactivation of MYC signaling confers resistance that can be reversed by the inhibition of MYC signaling. R-2HG also displays anti-tumor activity in glioma. Collectively, while R-2HG accumulated in IDH1/2 mutant cancers contributes to cancer initiation, our work demonstrates anti-tumor effects of 2HG in inhibiting proliferation/survival of FTO-high cancer cells via targeting FTO/m6A/MYC/CEBPA signaling.
Keywords: CEBPA; FTO; IDH mutation; MYC; N(6)-methyladenosine (m(6)A); R-2HG; S-2HG; glioma; leukemia.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
R-2HG Targets FTO to Increase m6A Levels and Suppress Tumor Growth.Cancer Discov. 2018 Feb;8(2):137. doi: 10.1158/2159-8290.CD-RW2017-240. Epub 2017 Dec 21. Cancer Discov. 2018. PMID: 29269348
References
-
- Boutzen H, Saland E, Larrue C, de Toni F, Gales L, Castelli FA, Cathebas M, Zaghdoudi S, Stuani L, Kaoma T, et al. Isocitrate dehydrogenase 1 mutations prime the all-trans retinoic acid myeloid differentiation pathway in acute myeloid leukemia. The Journal of experimental medicine. 2016;213:483–497. - PMC - PubMed
-
- Bralten LB, Kloosterhof NK, Balvers R, Sacchetti A, Lapre L, Lamfers M, Leenstra S, de Jonge H, Kros JM, Jansen EE, et al. IDH1 R132H decreases proliferation of glioma cell lines in vitro and in vivo. Annals of neurology. 2011;69:455–463. - PubMed
-
- Chou WC, Lei WC, Ko BS, Hou HA, Chen CY, Tang JL, Yao M, Tsay W, Wu SJ, Huang SY, et al. The prognostic impact and stability of Isocitrate dehydrogenase 2 mutation in adult patients with acute myeloid leukemia. Leukemia. 2011;25:246–253. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- RM1 HG008935/HG/NHGRI NIH HHS/United States
- R50 CA211404/CA/NCI NIH HHS/United States
- R01 CA182528/CA/NCI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R01 GM071440/GM/NIGMS NIH HHS/United States
- R01 CA214965/CA/NCI NIH HHS/United States
- R01 NS089815/NS/NINDS NIH HHS/United States
- R01 CA168815/CA/NCI NIH HHS/United States
- R21 NS100077/NS/NINDS NIH HHS/United States
- P30 CA033572/CA/NCI NIH HHS/United States
- R01 CA243386/CA/NCI NIH HHS/United States
- R01 CA178454/CA/NCI NIH HHS/United States
- R01 CA211614/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
