

Cullin 3-Based Ubiquitin Ligases as Master Regulators of Mammalian Cell Differentiation
- PMID: 29249570
- PMCID: PMC5801050
- DOI: 10.1016/j.tibs.2017.11.010
Cullin 3-Based Ubiquitin Ligases as Master Regulators of Mammalian Cell Differentiation
Abstract
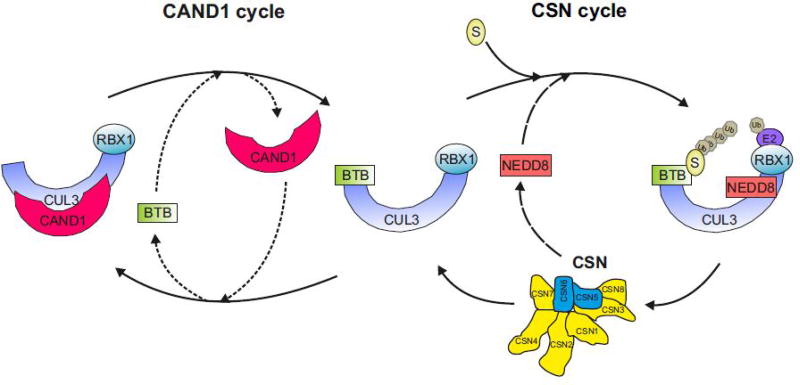
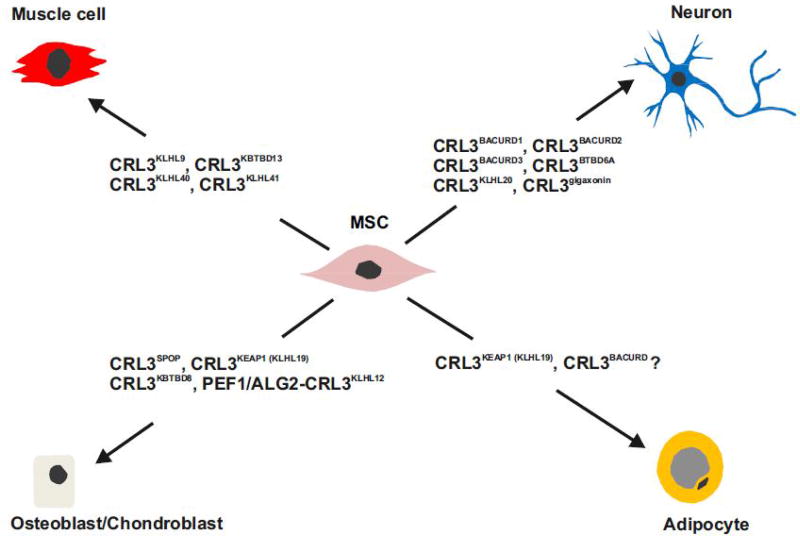
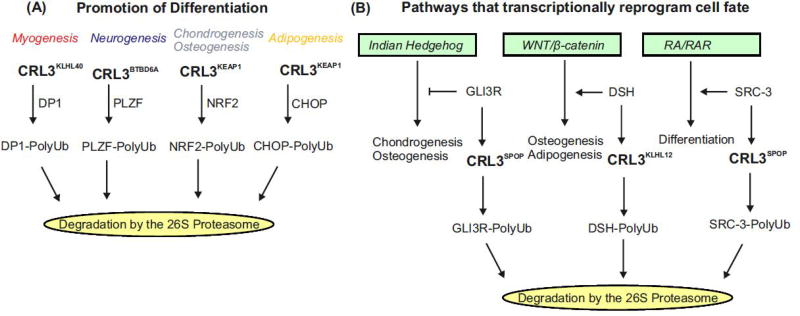
Specificity of the ubiquitin proteasome system is controlled by ubiquitin E3 ligases, including their major representatives, the multisubunit cullin-RING ubiquitin (Ub) ligases (CRLs). More than 200 different CRLs are divided into seven families according to their cullin scaffolding proteins (CUL1-7) around which they are assembled. Research over two decades has revealed that different CRL families are specialized to fulfill specific cellular functions. Whereas many CUL1-based CRLs (CRL1s) ubiquitylate cell cycle regulators, CRL4 complexes often associate with chromatin to control DNA metabolism. Based on studies about differentiation programs of mesenchymal stem cells (MSCs), including myogenesis, neurogenesis, chondrogenesis, osteogenesis and adipogenesis, we propose here that CRL3 complexes evolved to fulfill a pivotal role in mammalian cell differentiation.
Keywords: BTB proteins; Cullin 3; cullin-RING-ubiquitin ligases; cytoskeleton; differentiation; mesenchymal stem cells.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures

Similar articles
-
Rotavirus NSP1 Associates with Components of the Cullin RING Ligase Family of E3 Ubiquitin Ligases.J Virol. 2016 Jun 10;90(13):6036-48. doi: 10.1128/JVI.00704-16. Print 2016 Jul 1. J Virol. 2016. PMID: 27099313 Free PMC article.
-
Cullin-RING E3 Ubiquitin Ligases: Bridges to Destruction.Subcell Biochem. 2017;83:323-347. doi: 10.1007/978-3-319-46503-6_12. Subcell Biochem. 2017. PMID: 28271482 Free PMC article. Review.
-
Cullin 5-RING E3 ubiquitin ligases, new therapeutic targets?Biochimie. 2016 Mar;122:339-47. doi: 10.1016/j.biochi.2015.08.003. Epub 2015 Aug 4. Biochimie. 2016. PMID: 26253693 Review.
-
CAND1-dependent control of cullin 1-RING Ub ligases is essential for adipogenesis.Biochim Biophys Acta. 2013 May;1833(5):1078-84. doi: 10.1016/j.bbamcr.2013.01.005. Epub 2013 Jan 14. Biochim Biophys Acta. 2013. PMID: 23328082
-
CSN-CRL Complexes: New Regulators of Adipogenesis.Biomolecules. 2025 Mar 5;15(3):372. doi: 10.3390/biom15030372. Biomolecules. 2025. PMID: 40149914 Free PMC article. Review.
Cited by
-
Molecular choreography of E1 enzymes in ubiquitin-like protein cascades: New insights into dynamics and specificity.J Biol Chem. 2025 Aug;301(8):110415. doi: 10.1016/j.jbc.2025.110415. Epub 2025 Jun 24. J Biol Chem. 2025. PMID: 40570956 Free PMC article. Review.
-
Vimentin Localization in the Zebrafish Oral Cavity: A Potential Role in Taste Buds Regeneration.Int J Mol Sci. 2023 Oct 26;24(21):15619. doi: 10.3390/ijms242115619. Int J Mol Sci. 2023. PMID: 37958598 Free PMC article.
-
TRIM31 promotes glioma proliferation and invasion through activating NF-κB pathway.Onco Targets Ther. 2019 Mar 27;12:2289-2297. doi: 10.2147/OTT.S183625. eCollection 2019. Onco Targets Ther. 2019. Retraction in: Onco Targets Ther. 2024 Aug 20;17:671-672. doi: 10.2147/OTT.S491281. PMID: 30988633 Free PMC article. Retracted.
-
SPT5 stabilization of promoter-proximal RNA polymerase II.Mol Cell. 2021 Nov 4;81(21):4413-4424.e5. doi: 10.1016/j.molcel.2021.08.006. Epub 2021 Sep 3. Mol Cell. 2021. PMID: 34480849 Free PMC article.
-
PAR-1 signaling on macrophages is required for effective in vivo delayed-type hypersensitivity responses.iScience. 2021 Jan 5;24(1):101981. doi: 10.1016/j.isci.2020.101981. eCollection 2021 Jan 22. iScience. 2021. PMID: 33458623 Free PMC article.
References
-
- Deshaies RJ, Joazeiro CA. RING domain E3 ubiquitin ligases. Annu Rev Biochem. 2009;78:399–434. - PubMed
-
- Pintard L, Willis JH, Willems A, Johnson JL, Srayko M, Kurz T, Glaser S, Mains PE, Tyers M, Bowerman B, Peter M. The BTB protein MEL-26 is a substrate-specific adaptor of the CUL-3 ubiquitin-ligase. Nature. 2003;425:311–316. - PubMed
-
- Teixeira LK, Reed SI. Ubiquitin ligases and cell cycle control. Annu Rev Biochem. 2013;82:387–414. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
