

Schizophrenia and depression, two poles of endocannabinoid system deregulation
- PMID: 29249810
- PMCID: PMC5802629
- DOI: 10.1038/s41398-017-0029-y
Schizophrenia and depression, two poles of endocannabinoid system deregulation
Abstract
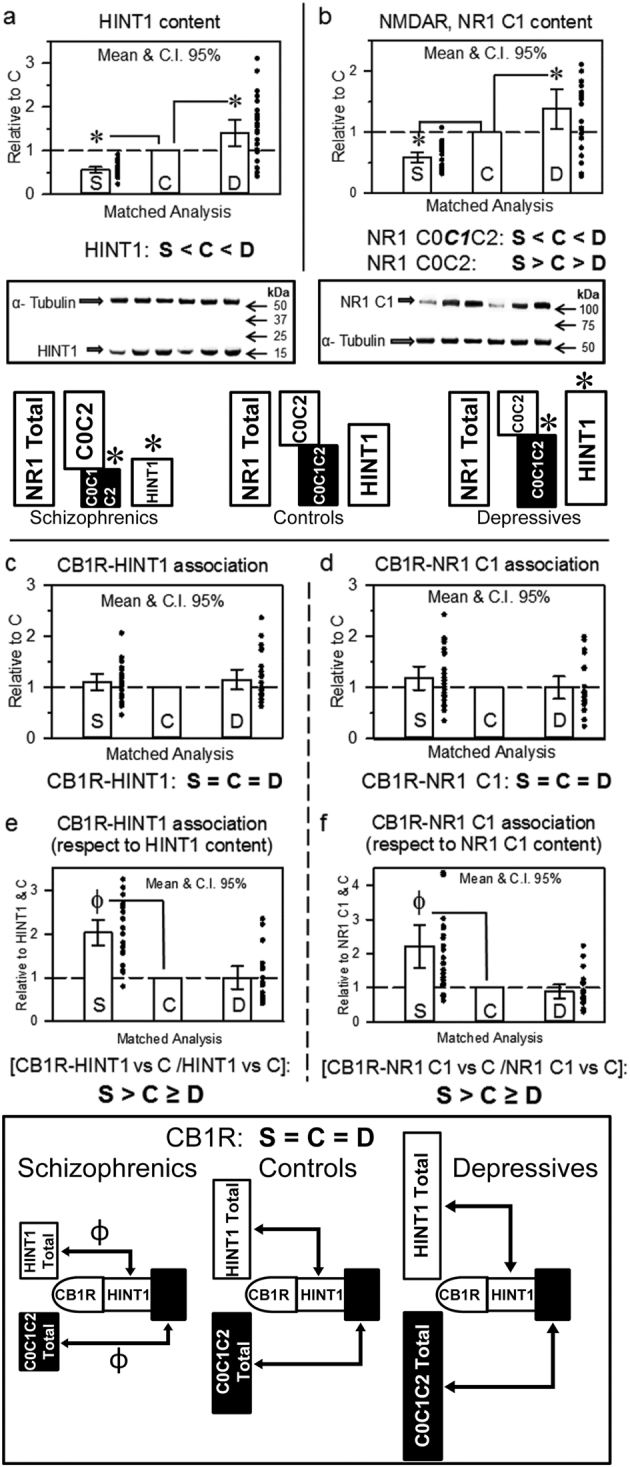
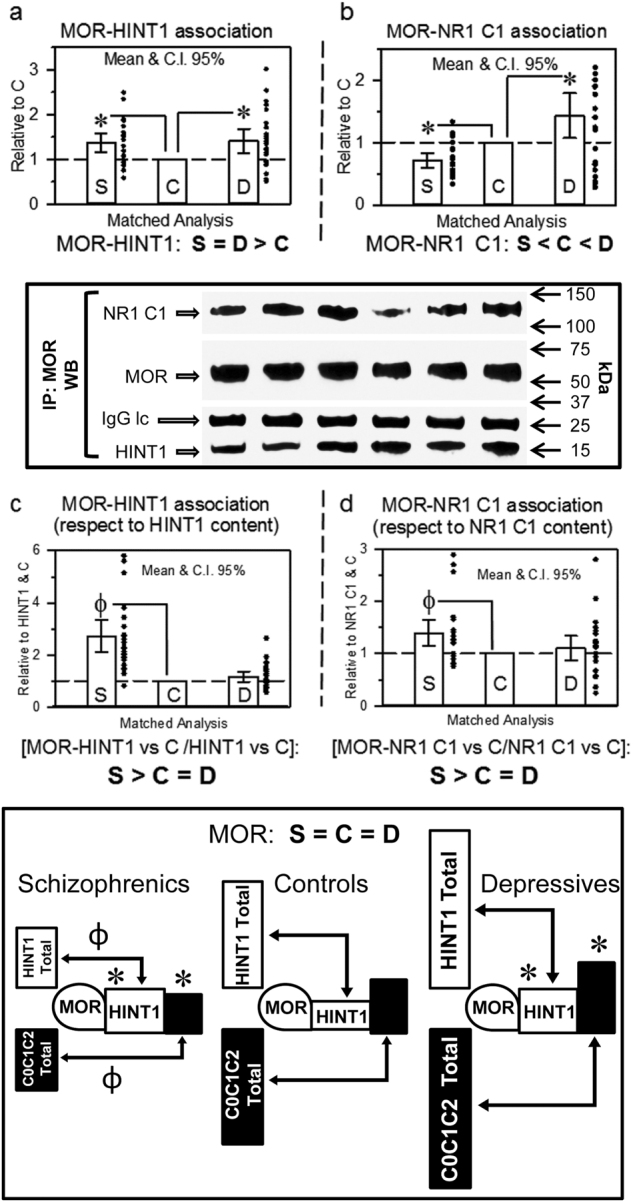
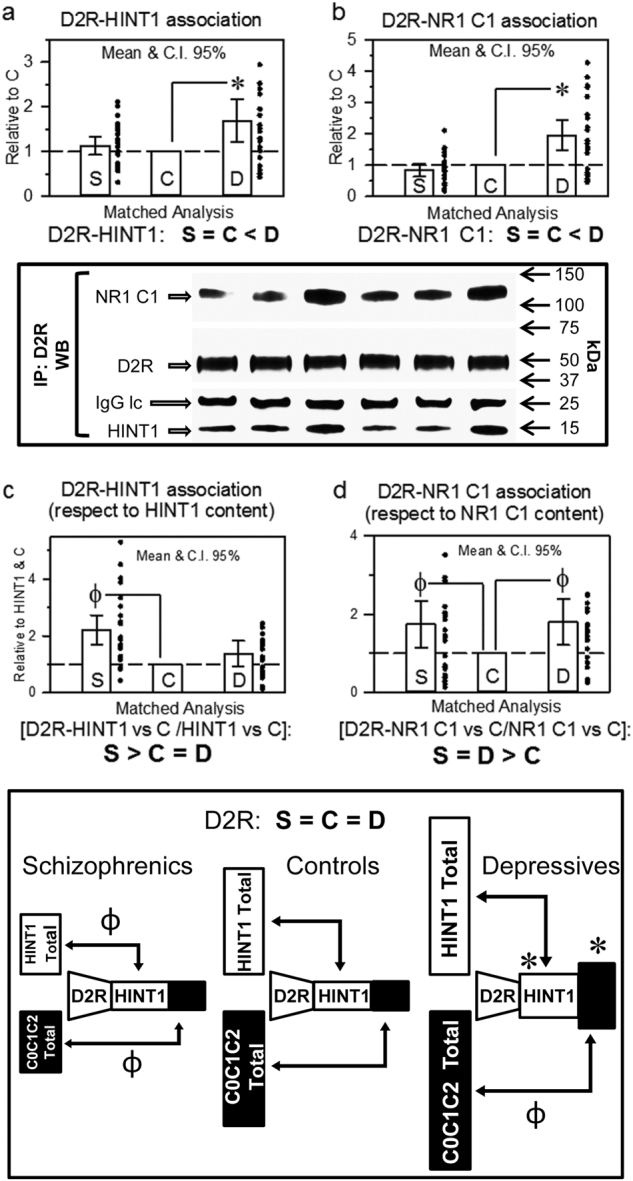
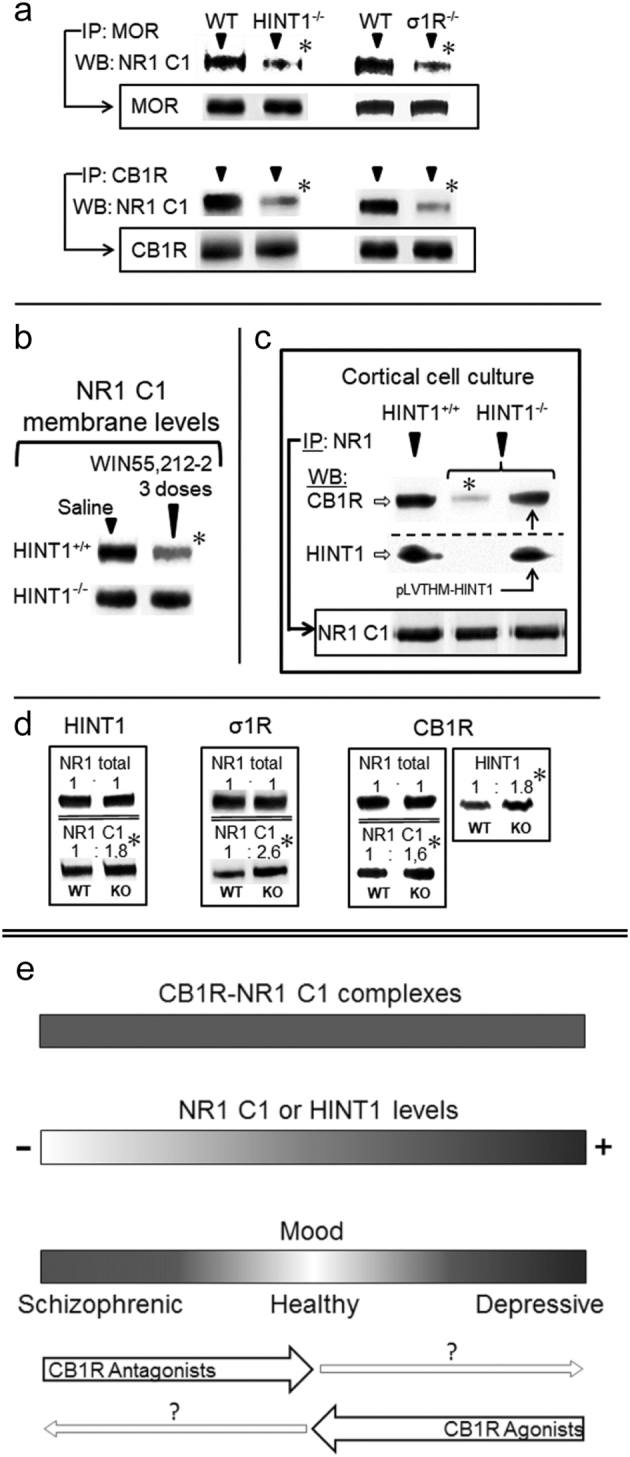
The activity of certain G protein-coupled receptors (GPCRs) and of glutamate N-Methyl-D-aspartate receptors (NMDARs) is altered in both schizophrenia and depression. Using postmortem prefrontal cortex samples from subjects with schizophrenia or depression, we observed a series of opposite changes in the expression of signaling proteins that have been implicated in the cross-talk between GPCRs and NMDARs. Thus, the levels of HINT1 proteins and NMDAR NR1 subunits carrying the C1 cytosolic segment were increased in depressives and decreased in schizophrenics, respect to matched controls. The differences in NR1 C1 subunits were compensated for via altered expression of NR1 subunits lacking the C1 segment; thus, the total number of NR1 subunits was comparable among the three groups. GPCRs influence the function of NR1 C1-containing NMDARs via PKC/Src, and thus, the association of mu-opioid and dopamine 2 receptors with NR1 C1 subunits was augmented in depressives and decreased in schizophrenics. However, the association of cannabinoid 1 receptors (CB1Rs) with NR1 C1 remained nearly constant. Endocannabinoids, via CB1Rs, control the presence of NR1 C1 subunits in the neural membrane. Thus, an altered endocannabinoid system may contribute to the pathophysiology of schizophrenia and depression by modifying the HINT1-NR1 C1/GPCR ratio, thereby altering GPCR-NMDAR cross-regulation.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
