

Leaf shedding as an anti-bacterial defense in Arabidopsis cauline leaves
- PMID: 29253890
- PMCID: PMC5749873
- DOI: 10.1371/journal.pgen.1007132
Leaf shedding as an anti-bacterial defense in Arabidopsis cauline leaves
Abstract
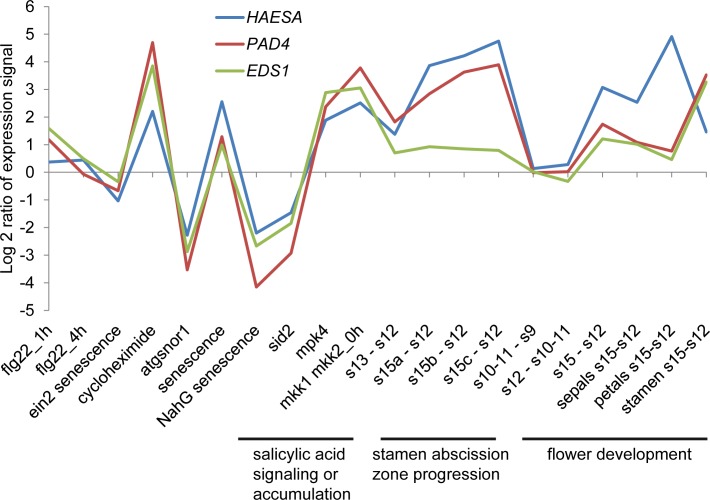
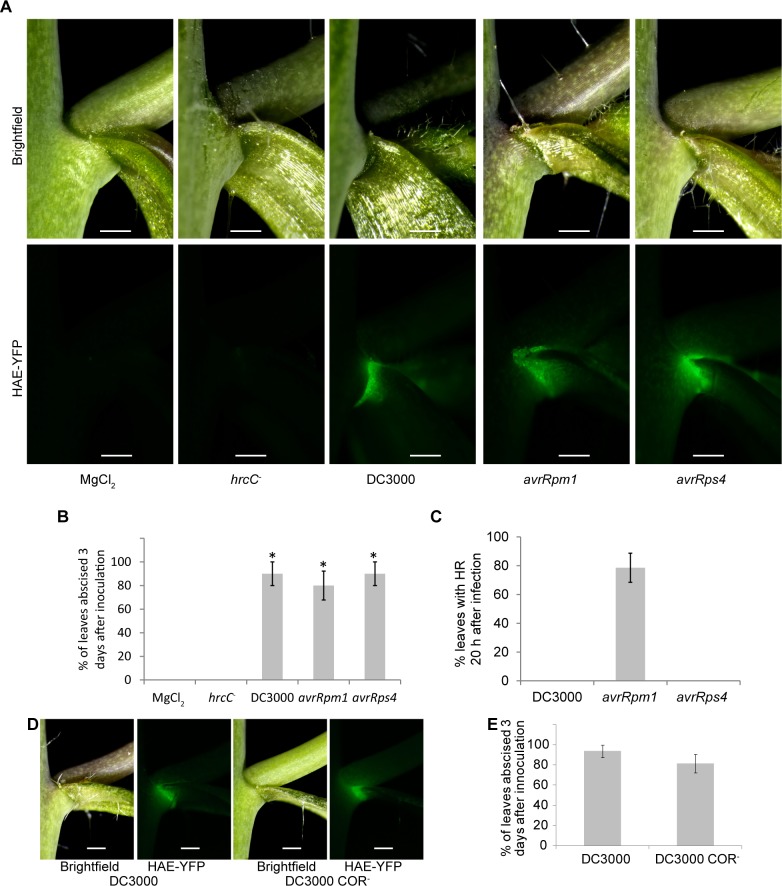
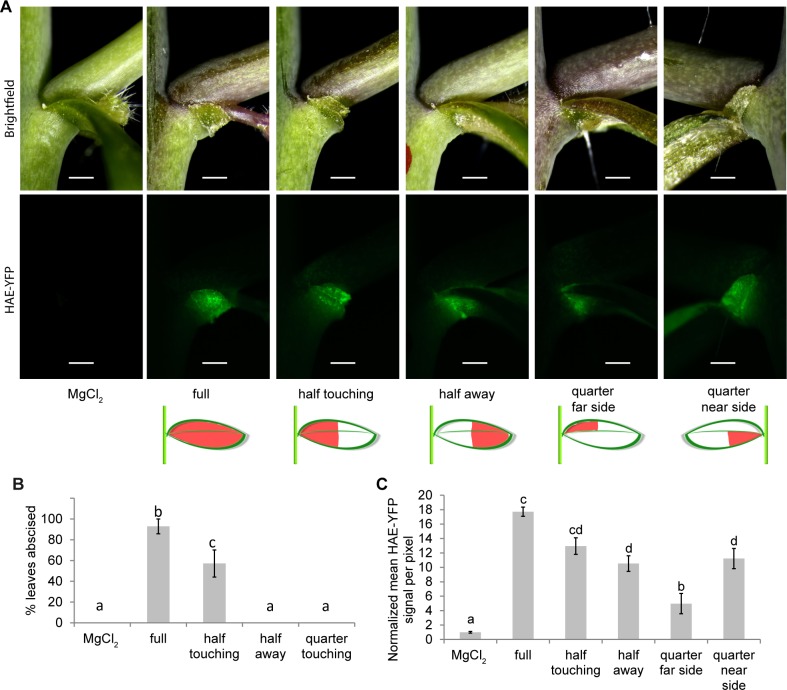
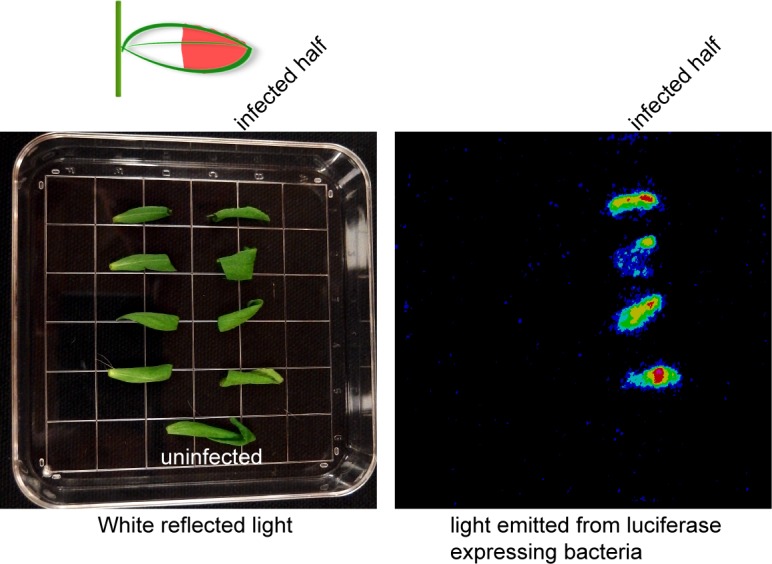
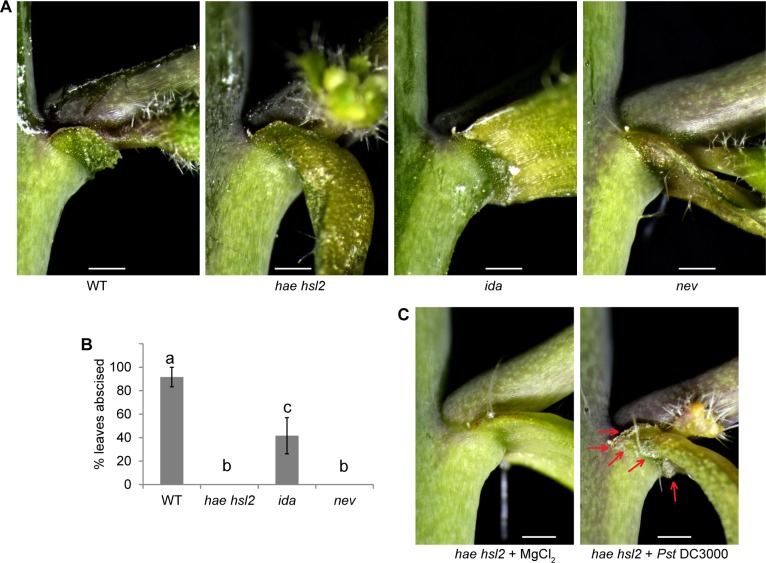
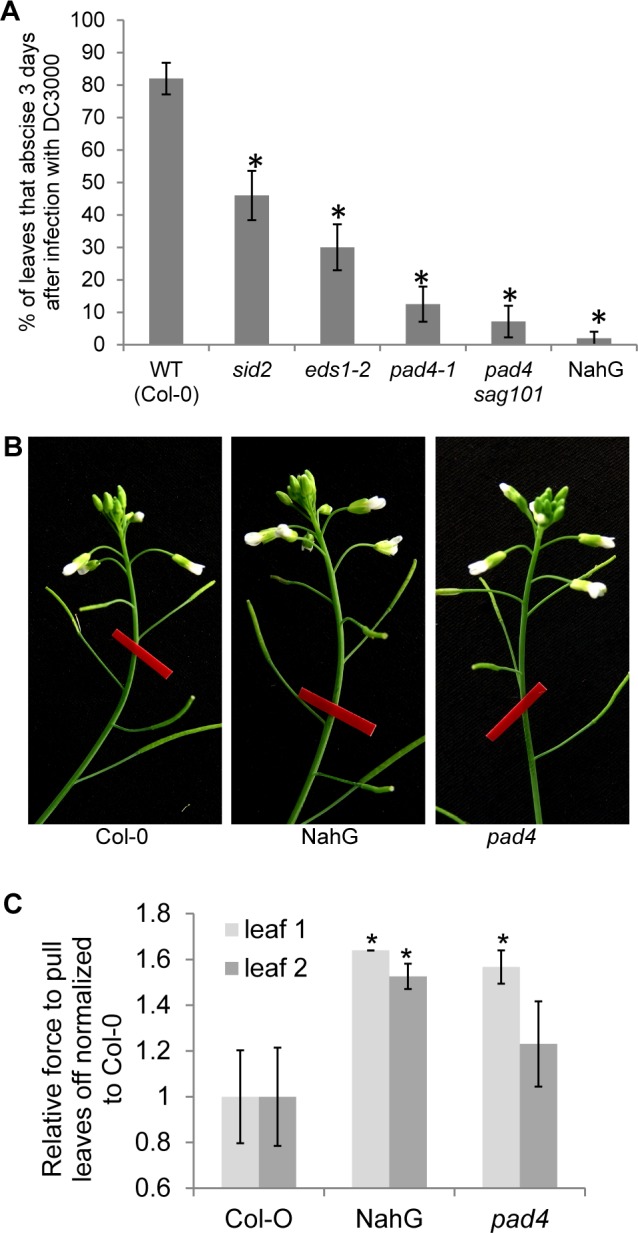
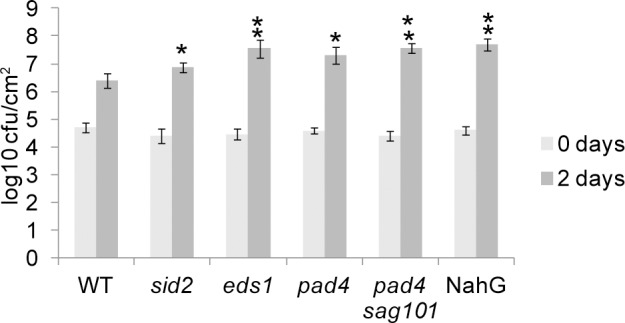
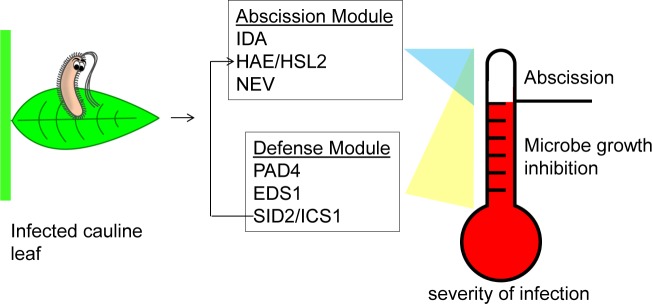
Plants utilize an innate immune system to protect themselves from disease. While many molecular components of plant innate immunity resemble the innate immunity of animals, plants also have evolved a number of truly unique defense mechanisms, particularly at the physiological level. Plant's flexible developmental program allows them the unique ability to simply produce new organs as needed, affording them the ability to replace damaged organs. Here we develop a system to study pathogen-triggered leaf abscission in Arabidopsis. Cauline leaves infected with the bacterial pathogen Pseudomonas syringae abscise as part of the defense mechanism. Pseudomonas syringae lacking a functional type III secretion system fail to elicit an abscission response, suggesting that the abscission response is a novel form of immunity triggered by effectors. HAESA/HAESA-like 2, INFLORESCENCE DEFICIENT IN ABSCISSION, and NEVERSHED are all required for pathogen-triggered abscission to occur. Additionally phytoalexin deficient 4, enhanced disease susceptibility 1, salicylic acid induction-deficient 2, and senescence-associated gene 101 plants with mutations in genes necessary for bacterial defense and salicylic acid signaling, and NahG transgenic plants with low levels of salicylic acid fail to abscise cauline leaves normally. Bacteria that physically contact abscission zones trigger a strong abscission response; however, long-distance signals are also sent from distal infected tissue to the abscission zone, alerting the abscission zone of looming danger. We propose a threshold model regulating cauline leaf defense where minor infections are handled by limiting bacterial growth, but when an infection is deemed out of control, cauline leaves are shed. Together with previous results, our findings suggest that salicylic acid may regulate both pathogen- and drought-triggered leaf abscission.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Abramovitch RB, Anderson JC, Martin GB. Bacterial elicitation and evasion of plant innate immunity. Nat Rev Mol Cell Biol. 2006;7: 601–611. doi: 10.1038/nrm1984 - DOI - PMC - PubMed
-
- Jones JDG, Dangl JL. The plant immune system. Nat Lond. 2006;444: 323–9. http://dx.doi.org/10.1038/nature05286 - DOI - PubMed
-
- Zipfel C. Pattern-recognition receptors in plant innate immunity. Curr Opin Immunol. 2008;20: 10–16. doi: 10.1016/j.coi.2007.11.003 - DOI - PubMed
-
- Melotto M, Underwood W, He SY. Role of Stomata in Plant Innate Immunity and Foliar Bacterial Diseases. Annu Rev Phytopathol. 2008;46: 101–122. doi: 10.1146/annurev.phyto.121107.104959 - DOI - PMC - PubMed
-
- Dodds PN, Rathjen JP. Plant immunity: towards an integrated view of plant–pathogen interactions. Nat Rev Genet. 2010;11: 539–548. doi: 10.1038/nrg2812 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
