

Mechanisms of gene regulation in human embryos and pluripotent stem cells
- PMID: 29254992
- PMCID: PMC5769625
- DOI: 10.1242/dev.157404
Mechanisms of gene regulation in human embryos and pluripotent stem cells
Abstract
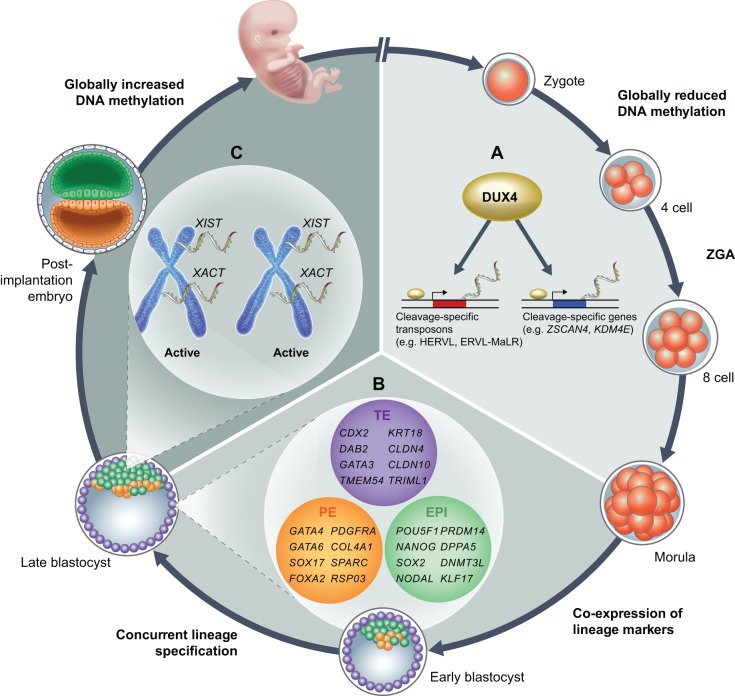
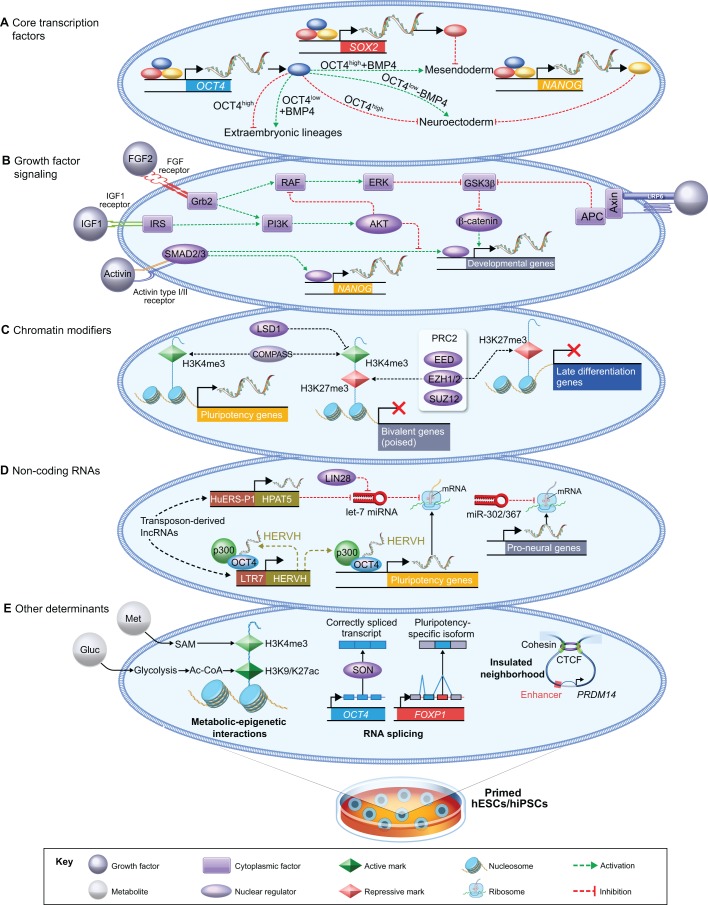
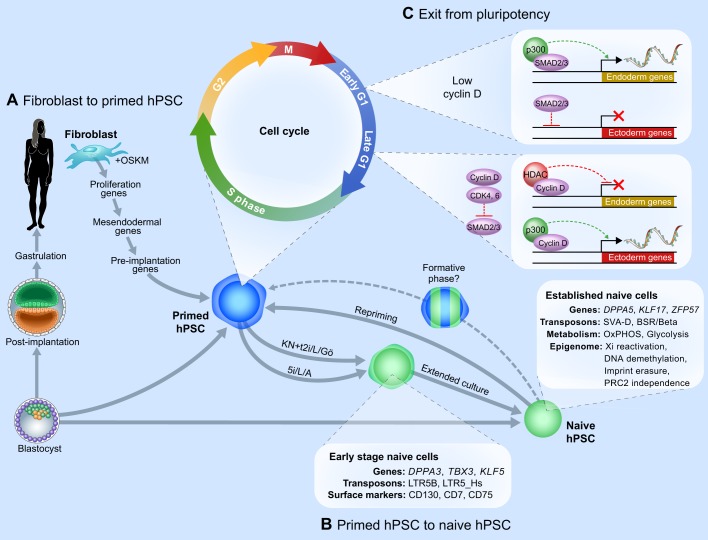
Pluripotent stem cells have broad utility in biomedical research and their molecular regulation has thus garnered substantial interest. While the principles that establish and regulate pluripotency have been well defined in the mouse, it has been difficult to extrapolate these insights to the human system due to species-specific differences and the distinct developmental identities of mouse versus human embryonic stem cells. In this Review, we examine genome-wide approaches to elucidate the regulatory principles of pluripotency in human embryos and stem cells, and highlight where differences exist in the regulation of pluripotency in mice and humans. We review recent insights into the nature of human pluripotent cells in vivo, obtained by the deep sequencing of pre-implantation embryos. We also present an integrated overview of the principal layers of global gene regulation in human pluripotent stem cells. Finally, we discuss the transcriptional and epigenomic remodeling events associated with cell fate transitions into and out of human pluripotency.
Keywords: Chromatin; Embryogenesis; Epigenetics; Pluripotency; Reprogramming; Transcription.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsR.J. is co-founder of Fate Therapeutics, Fulcrum Therapeutics and Omega Therapeutics.
Figures
References
-
- Anokye-Danso F., Trivedi C. M., Juhr D., Gupta M., Cui Z., Tian Y., Zhang Y., Yang W., Gruber P. J., Epstein J. A. et al. (2011). Highly efficient miRNA-mediated reprogramming of mouse and human somatic cells to pluripotency. Cell Stem Cell 8, 376-388. 10.1016/j.stem.2011.03.001 - DOI - PMC - PubMed
-
- Bar M., Wyman S. K., Fritz B. R., Qi J., Garg K. S., Parkin R. K., Kroh E. M., Bendoraite A., Mitchell P. S., Nelson A. M. et al. (2008). MicroRNA discovery and profiling in human embryonic stem cells by deep sequencing of small RNA libraries. Stem Cells 26, 2496-2505. 10.1634/stemcells.2008-0356 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
