

Functional characterization of the circadian clock in the Antarctic krill, Euphausia superba
- PMID: 29255161
- PMCID: PMC5735174
- DOI: 10.1038/s41598-017-18009-2
Functional characterization of the circadian clock in the Antarctic krill, Euphausia superba
Abstract
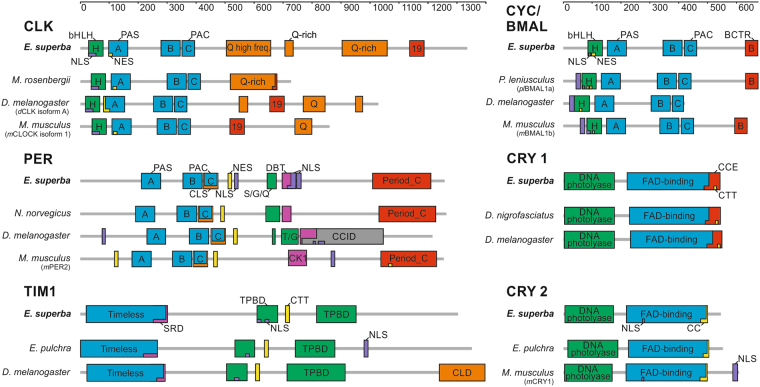
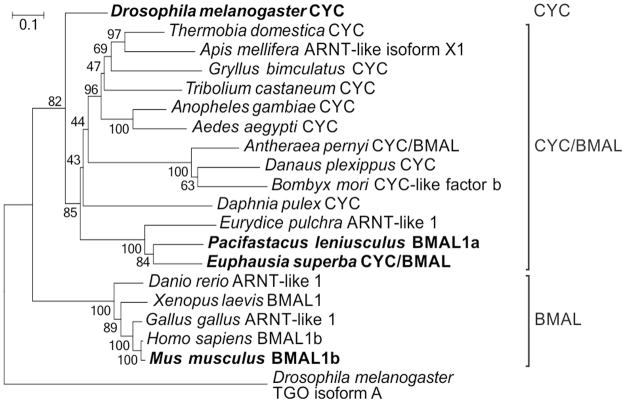
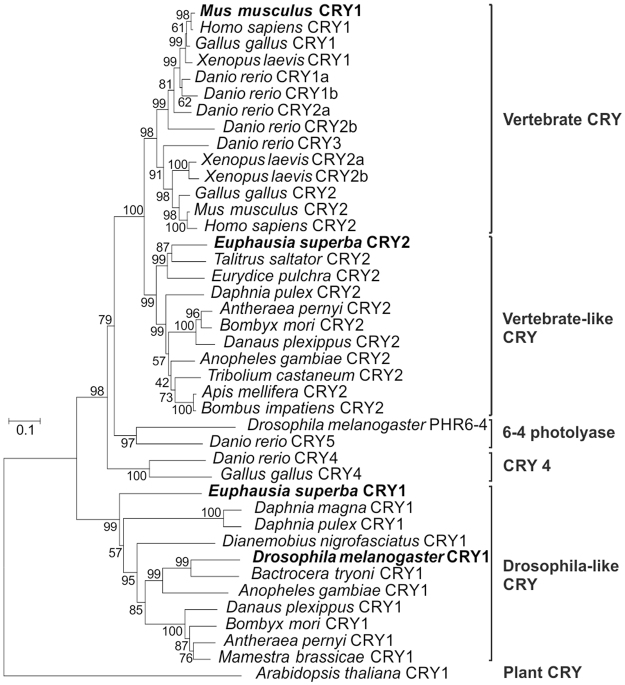
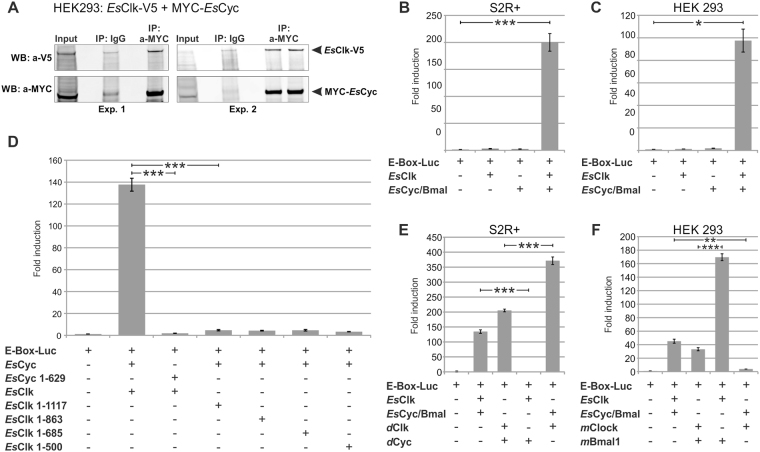
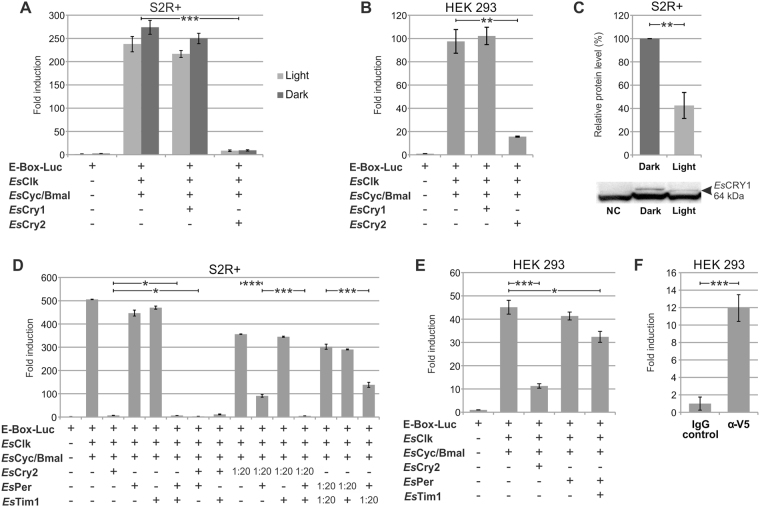
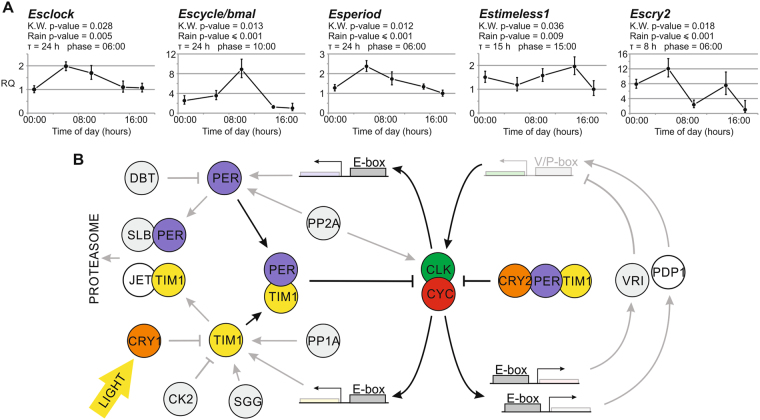
Antarctic krill (Euphausia superba) is a key species in Southern Ocean ecosystem where it plays a central role in the Antarctic food web. Available information supports the existence of an endogenous timing system in krill enabling it to synchronize metabolism and behavior with an environment characterized by extreme seasonal changes in terms of day length, food availability, and surface ice extent. A screening of our transcriptome database "KrillDB" allowed us to identify the putative orthologues of 20 circadian clock components. Mapping of conserved domains and phylogenetic analyses strongly supported annotations of the identified sequences. Luciferase assays and co-immunoprecipitation experiments allowed us to define the role of the main clock components. Our findings provide an overall picture of the molecular mechanisms underlying the functioning of the endogenous circadian clock in the Antarctic krill and shed light on their evolution throughout crustaceans speciation. Interestingly, the core clock machinery shows both mammalian and insect features that presumably contribute to an evolutionary strategy to cope with polar environment's challenges. Moreover, despite the extreme variability characterizing the Antarctic seasonal day length, the conserved light mediated degradation of the photoreceptor EsCRY1 suggests a persisting pivotal role of light as a Zeitgeber.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Marr, J. W. S. The natural history and geography of the Antarctic krill (Euphausia superba Dana). Discovery reports32 (1962).
-
- Croxall J, Reid K, Prince P. Diet, provisioning and productivity responses of marine predators to differences in availability of Antarctic krill. Mar. Ecol. Prog. Ser. 1999;177:115–131. doi: 10.3354/meps177115. - DOI
-
- Atkinson A, Siegel V, Pakhomov EA, Jessopp MJ, Loeb V. A re-appraisal of the total biomass and annual production of Antarctic krill. Deep Sea Res. Part I Oceanogr. Res. Pap. 2009;56:727–740. doi: 10.1016/j.dsr.2008.12.007. - DOI
-
- Nicol S, Foster J, Kawaguchi S. The fishery for Antarctic krill - recent developments. Fish Fish. 2012;13:30–40. doi: 10.1111/j.1467-2979.2011.00406.x. - DOI
-
- Ducklow H, et al. West Antarctic Peninsula: an ice-dependent coastal marine ecosystem in transition. Oceanography. 2013;26:190–203. doi: 10.5670/oceanog.2013.62. - DOI
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
