

Genomic features of bacterial adaptation to plants
- PMID: 29255260
- PMCID: PMC5957079
- DOI: 10.1038/s41588-017-0012-9
Genomic features of bacterial adaptation to plants
Abstract
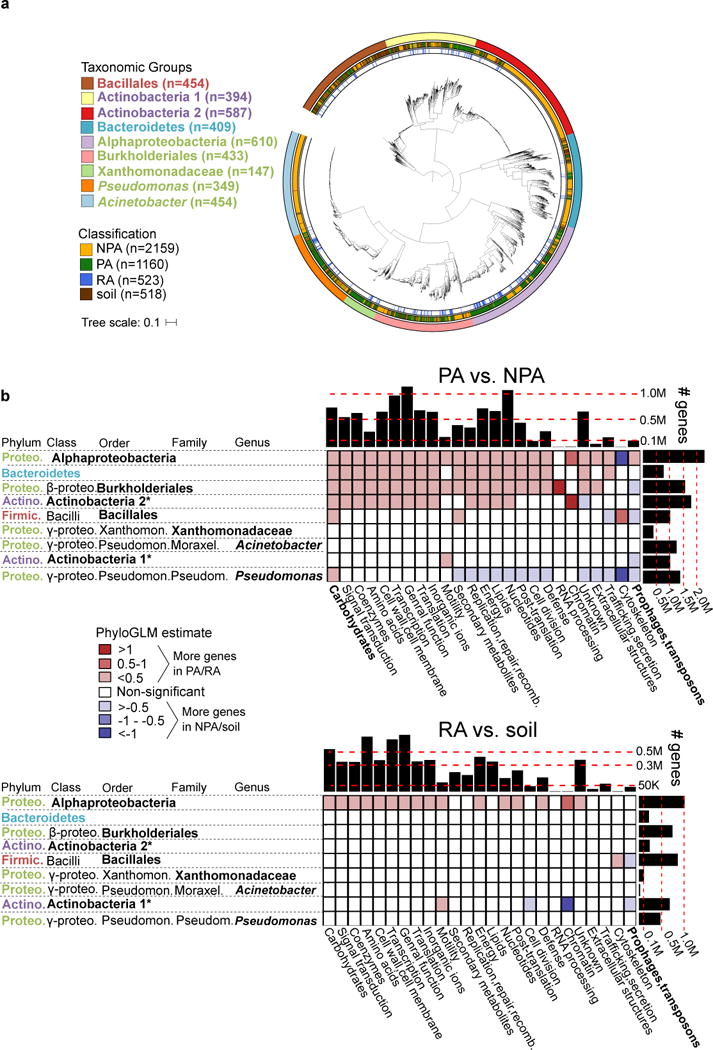
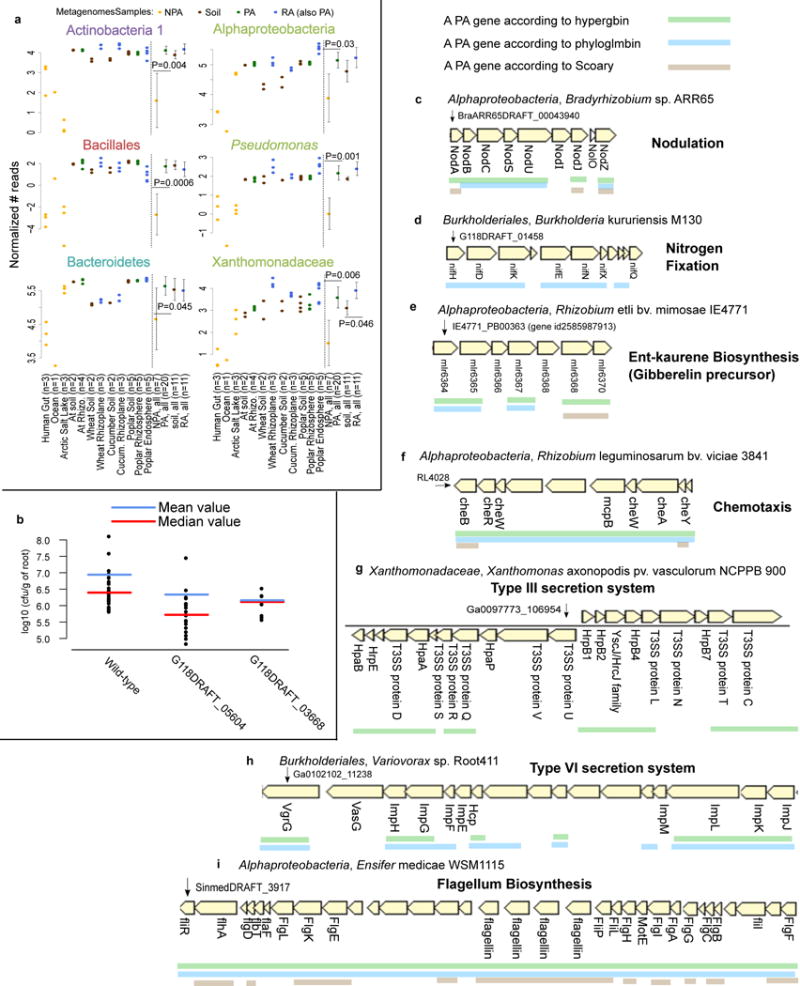
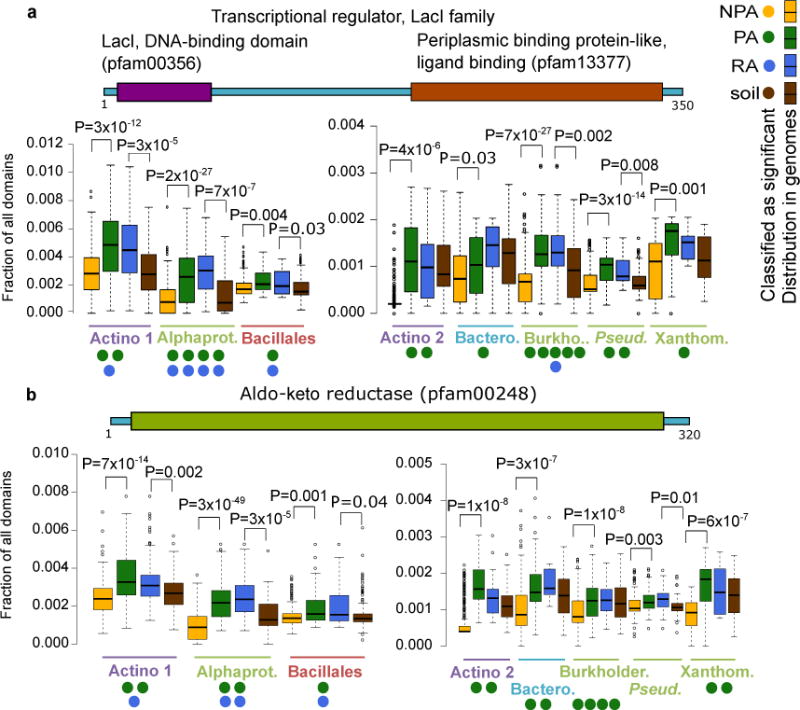
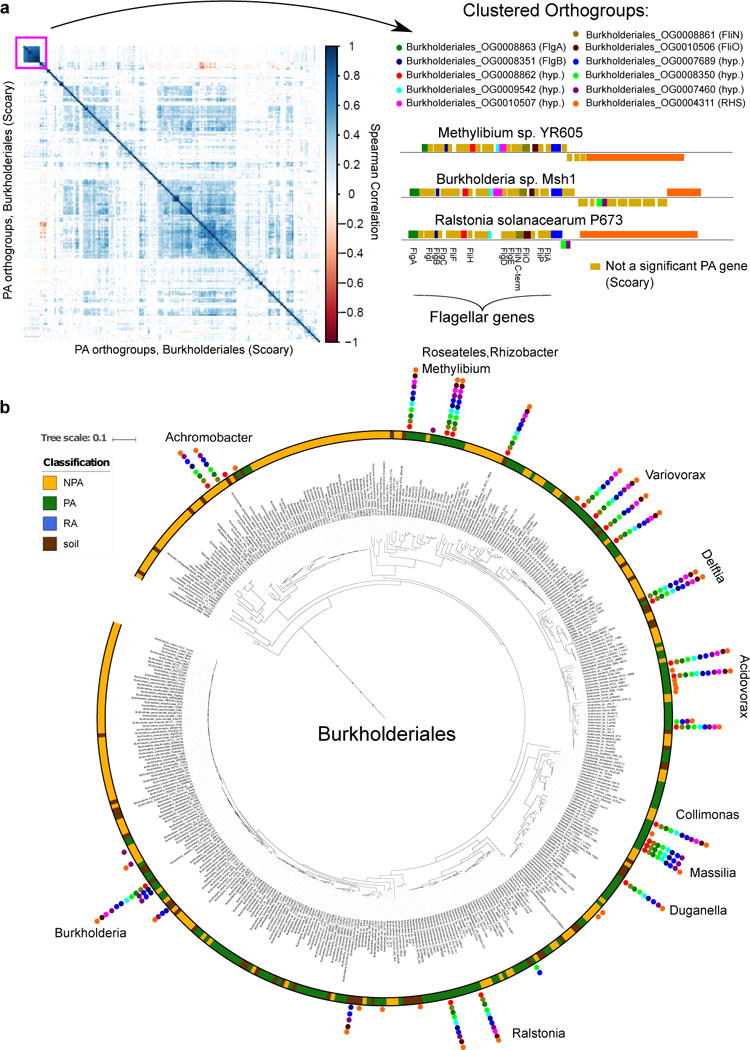
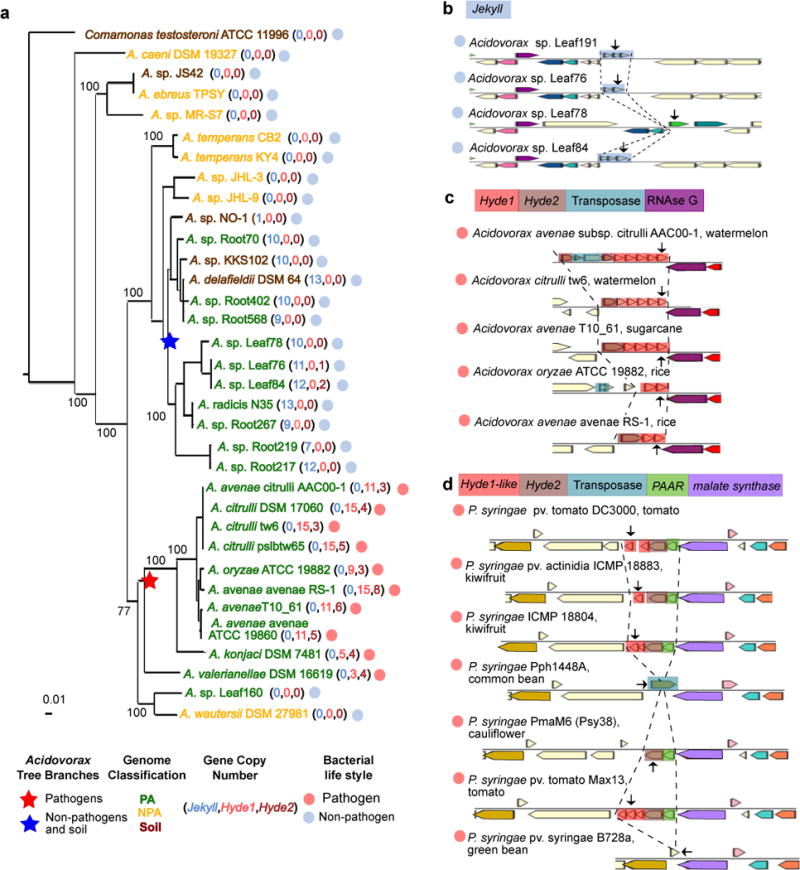
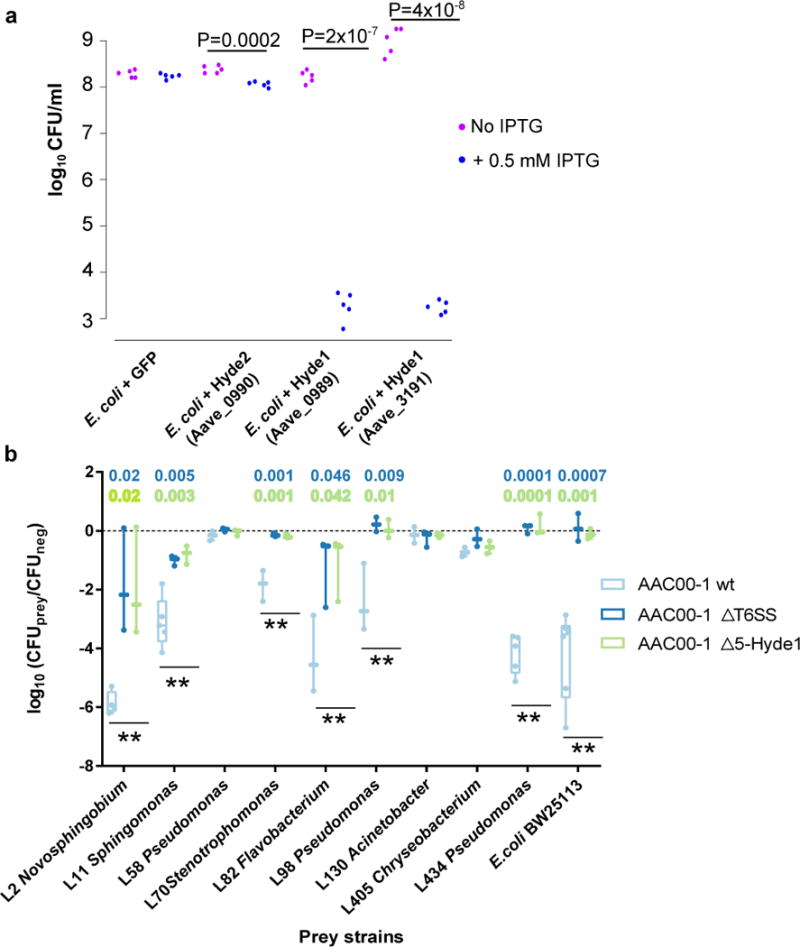
Plants intimately associate with diverse bacteria. Plant-associated bacteria have ostensibly evolved genes that enable them to adapt to plant environments. However, the identities of such genes are mostly unknown, and their functions are poorly characterized. We sequenced 484 genomes of bacterial isolates from roots of Brassicaceae, poplar, and maize. We then compared 3,837 bacterial genomes to identify thousands of plant-associated gene clusters. Genomes of plant-associated bacteria encode more carbohydrate metabolism functions and fewer mobile elements than related non-plant-associated genomes do. We experimentally validated candidates from two sets of plant-associated genes: one involved in plant colonization, and the other serving in microbe-microbe competition between plant-associated bacteria. We also identified 64 plant-associated protein domains that potentially mimic plant domains; some are shared with plant-associated fungi and oomycetes. This work expands the genome-based understanding of plant-microbe interactions and provides potential leads for efficient and sustainable agriculture through microbiome engineering.
Figures

Comment in
-
Bacterial genomics of plant adaptation.Nat Genet. 2018 Jan;50(1):2-4. doi: 10.1038/s41588-017-0019-2. Nat Genet. 2018. PMID: 29273805 No abstract available.
-
Adapt to plants.Nat Plants. 2018 Feb;4(2):60. doi: 10.1038/s41477-018-0110-4. Nat Plants. 2018. PMID: 29379147 No abstract available.
