

A mixed-kinetic model describes unloaded velocities of smooth, skeletal, and cardiac muscle myosin filaments in vitro
- PMID: 29255801
- PMCID: PMC5733112
- DOI: 10.1126/sciadv.aao2267
A mixed-kinetic model describes unloaded velocities of smooth, skeletal, and cardiac muscle myosin filaments in vitro
Abstract
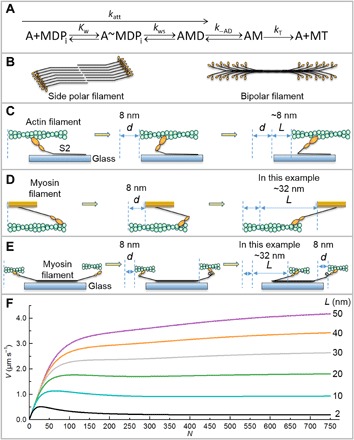
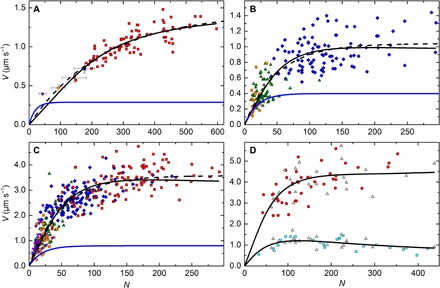
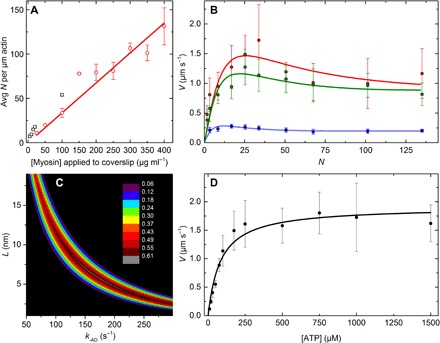
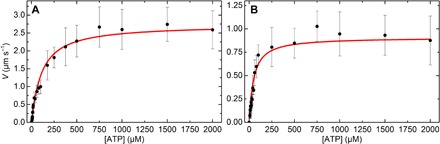
In vitro motility assays, where purified myosin and actin move relative to one another, are used to better understand the mechanochemistry of the actomyosin adenosine triphosphatase (ATPase) cycle. We examined the relationship between the relative velocity (V) of actin and myosin and the number of available myosin heads (N) or [ATP] for smooth (SMM), skeletal (SKM), and cardiac (CMM) muscle myosin filaments moving over actin as well as V from actin filaments moving over a bed of monomeric SKM. These data do not fit well to a widely accepted model that predicts that V is limited by myosin detachment from actin (d/ton), where d equals step size and ton equals time a myosin head remains attached to actin. To account for these data, we have developed a mixed-kinetic model where V is influenced by both attachment and detachment kinetics. The relative contributions at a given V vary with the probability that a head will remain attached to actin long enough to reach the end of its flexible S2 tether. Detachment kinetics are affected by L/ton, where L is related to the tether length. We show that L is relatively long for SMM, SKM, and CMM filaments (59 ± 3 nm, 22 ± 9 nm, and 22 ± 2 nm, respectively). In contrast, L is shorter (8 ± 3 nm) when myosin monomers are attached to a surface. This suggests that the behavior of the S2 domain may be an important mechanical feature of myosin filaments that influences unloaded shortening velocities of muscle.
Figures
References
-
- Yanagida T., Nakase M., Nishiyama K., Oosawa F., Direct observation of motion of single F-actin filaments in the presence of myosin. Nature 307, 58–60 (1984). - PubMed
-
- Toyoshima Y. Y., Kron S. J., McNally E. M., Niebling K. R., Toyoshima C., Spudich J. A., Myosin subfragment-1 is sufficient to move actin filaments in vitro. Nature 328, 536–539 (1987). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
