

Stress Granule-Inducing Eukaryotic Translation Initiation Factor 4A Inhibitors Block Influenza A Virus Replication
- PMID: 29258238
- PMCID: PMC5744162
- DOI: 10.3390/v9120388
Stress Granule-Inducing Eukaryotic Translation Initiation Factor 4A Inhibitors Block Influenza A Virus Replication
Abstract
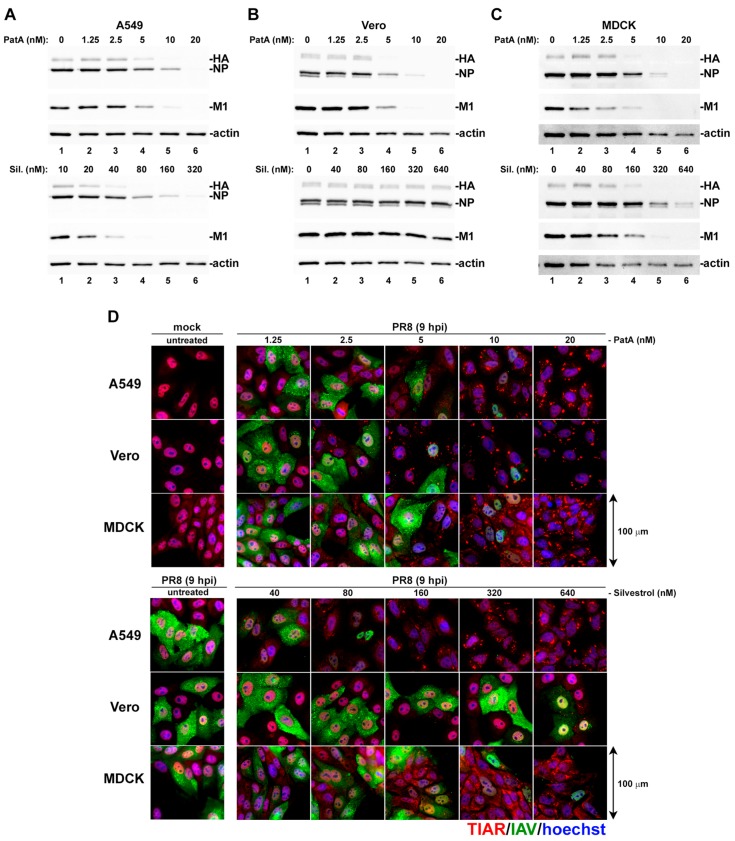
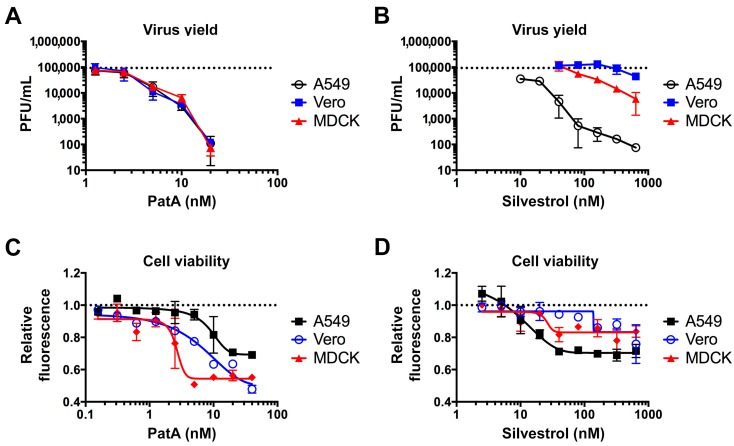
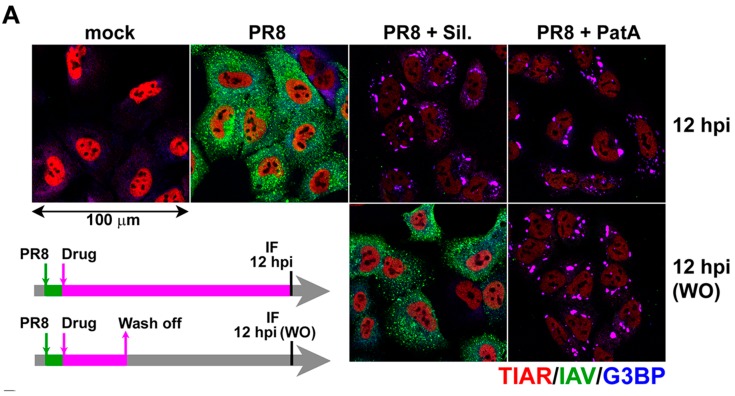
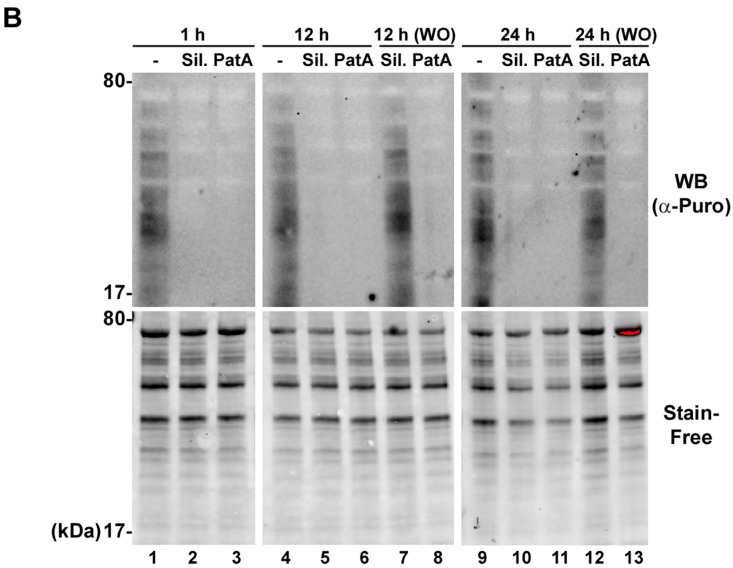
Eukaryotic translation initiation factor 4A (eIF4A) is a helicase that facilitates assembly of the translation preinitiation complex by unwinding structured mRNA 5' untranslated regions. Pateamine A (PatA) and silvestrol are natural products that disrupt eIF4A function and arrest translation, thereby triggering the formation of cytoplasmic aggregates of stalled preinitiation complexes known as stress granules (SGs). Here we examined the effects of eIF4A inhibition by PatA and silvestrol on influenza A virus (IAV) protein synthesis and replication in cell culture. Treatment of infected cells with either PatA or silvestrol at early times post-infection resulted in SG formation, arrest of viral protein synthesis and failure to replicate the viral genome. PatA, which irreversibly binds to eIF4A, sustained long-term blockade of IAV replication following drug withdrawal, and inhibited IAV replication at concentrations that had minimal cytotoxicity. By contrast, the antiviral effects of silvestrol were fully reversible; drug withdrawal caused rapid SG dissolution and resumption of viral protein synthesis. IAV inhibition by silvestrol was invariably associated with cytotoxicity. PatA blocked replication of genetically divergent IAV strains, suggesting common dependence on host eIF4A activity. This study demonstrates that the core host protein synthesis machinery can be targeted to block viral replication.
Keywords: eIF4A helicase; influenza A virus; pateamine A; silvestrol; stress granule; translation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
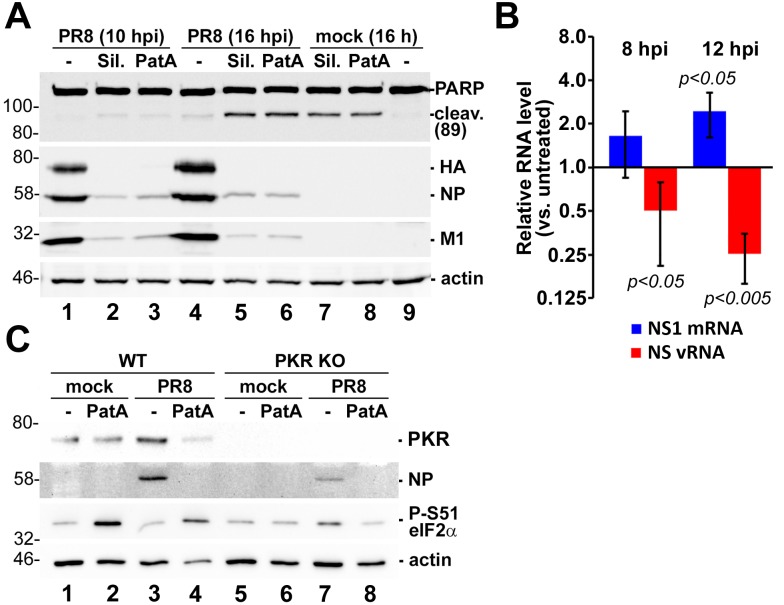
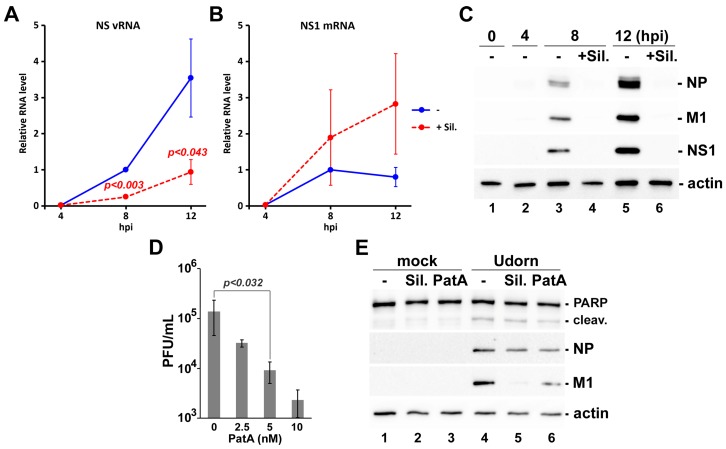
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
