

Permeating disciplines: Overcoming barriers between molecular simulations and classical structure-function approaches in biological ion transport
- PMID: 29258839
- PMCID: PMC6317864
- DOI: 10.1016/j.bbamem.2017.12.013
Permeating disciplines: Overcoming barriers between molecular simulations and classical structure-function approaches in biological ion transport
Abstract
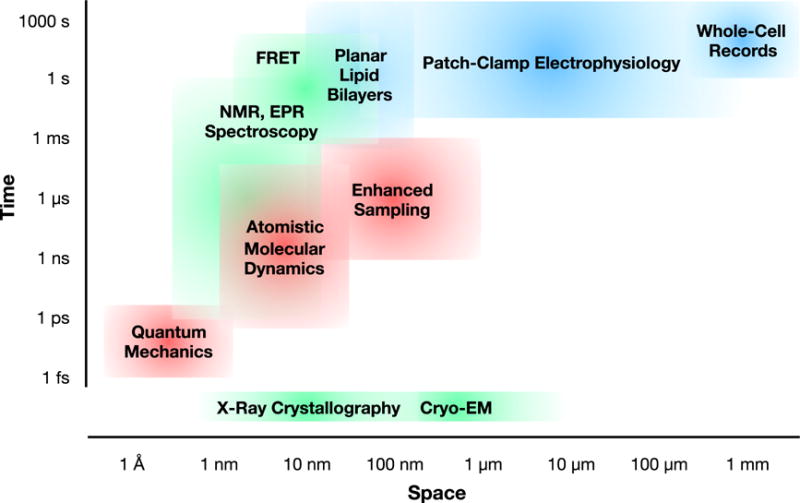
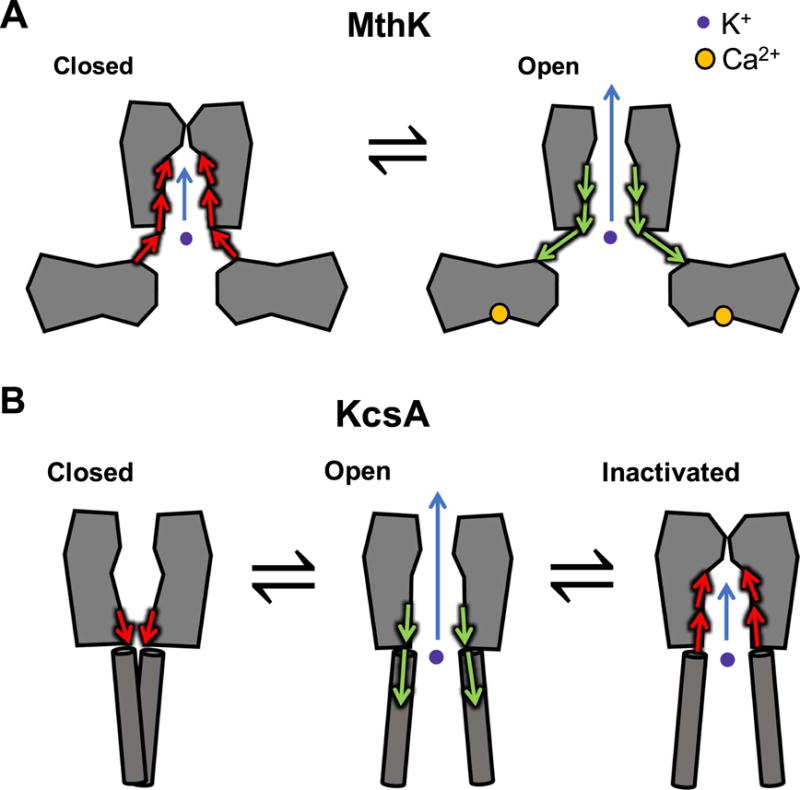
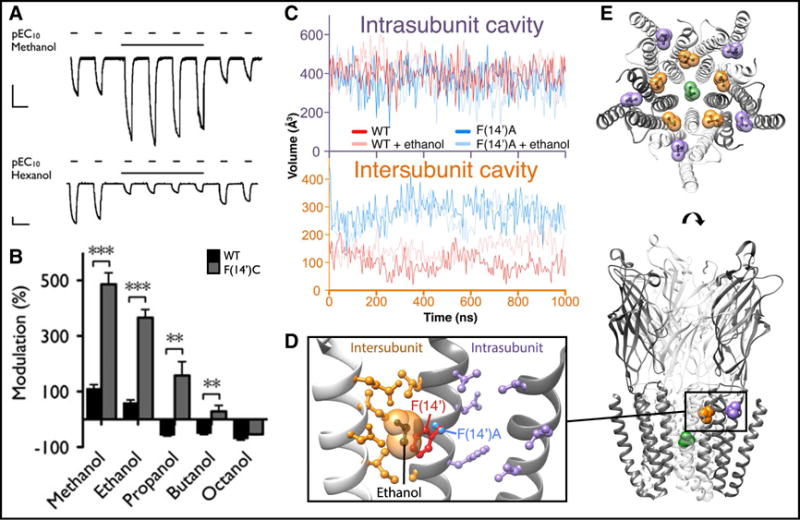
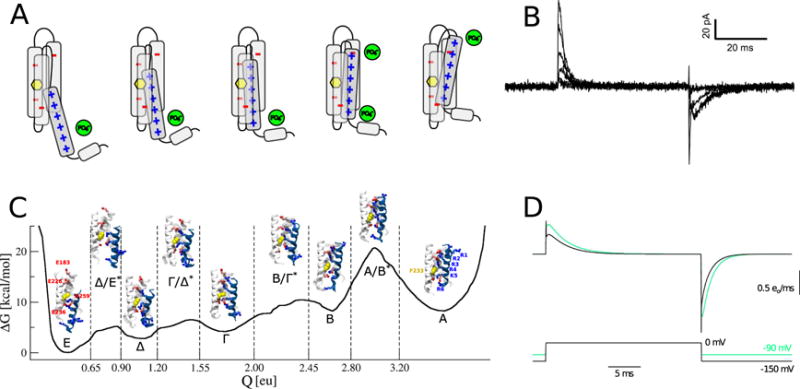
Ion translocation across biological barriers is a fundamental requirement for life. In many cases, controlling this process-for example with neuroactive drugs-demands an understanding of rapid and reversible structural changes in membrane-embedded proteins, including ion channels and transporters. Classical approaches to electrophysiology and structural biology have provided valuable insights into several such proteins over macroscopic, often discontinuous scales of space and time. Integrating these observations into meaningful mechanistic models now relies increasingly on computational methods, particularly molecular dynamics simulations, while surfacing important challenges in data management and conceptual alignment. Here, we seek to provide contemporary context, concrete examples, and a look to the future for bridging disciplinary gaps in biological ion transport. This article is part of a Special Issue entitled: Beyond the Structure-Function Horizon of Membrane Proteins edited by Ute Hellmich, Rupak Doshi and Benjamin McIlwain.
Keywords: Electrophysiology; Ion channel; Ion transport; Kinetic modeling; Molecular dynamics; Structural biology.
Copyright © 2017 Elsevier B.V. All rights reserved.
Figures
Similar articles
-
Membrane transporter research in times of countless structures.Biochim Biophys Acta Biomembr. 2018 Apr;1860(4):804-808. doi: 10.1016/j.bbamem.2017.08.009. Epub 2017 Aug 31. Biochim Biophys Acta Biomembr. 2018. PMID: 28867210 Review.
-
Insights into the function of ion channels by computational electrophysiology simulations.Biochim Biophys Acta. 2016 Jul;1858(7 Pt B):1741-52. doi: 10.1016/j.bbamem.2016.02.006. Epub 2016 Feb 10. Biochim Biophys Acta. 2016. PMID: 26874204 Review.
-
Computational studies of transport in ion channels using metadynamics.Biochim Biophys Acta. 2016 Jul;1858(7 Pt B):1733-40. doi: 10.1016/j.bbamem.2016.02.015. Epub 2016 Feb 15. Biochim Biophys Acta. 2016. PMID: 26891818 Review.
-
Simulations of outer membrane channels and their permeability.Biochim Biophys Acta. 2016 Jul;1858(7 Pt B):1760-71. doi: 10.1016/j.bbamem.2015.12.020. Epub 2015 Dec 23. Biochim Biophys Acta. 2016. PMID: 26721326 Review.
-
Molecular dynamics simulations of membrane channels and transporters.Curr Opin Struct Biol. 2009 Apr;19(2):128-37. doi: 10.1016/j.sbi.2009.02.011. Epub 2009 Apr 1. Curr Opin Struct Biol. 2009. PMID: 19345092 Free PMC article. Review.
Cited by
-
Photopharmacology of Ion Channels through the Light of the Computational Microscope.Int J Mol Sci. 2021 Nov 8;22(21):12072. doi: 10.3390/ijms222112072. Int J Mol Sci. 2021. PMID: 34769504 Free PMC article. Review.
-
Roles for Countercharge in the Voltage Sensor Domain of Ion Channels.Front Pharmacol. 2020 Feb 28;11:160. doi: 10.3389/fphar.2020.00160. eCollection 2020. Front Pharmacol. 2020. PMID: 32180723 Free PMC article. Review.
-
Molecular dynamics: a powerful tool for studying the medicinal chemistry of ion channel modulators.RSC Med Chem. 2021 Jul 22;12(9):1503-1518. doi: 10.1039/d1md00140j. eCollection 2021 Sep 23. RSC Med Chem. 2021. PMID: 34671734 Free PMC article. Review.
-
The Structural Basis of IKs Ion-Channel Activation: Mechanistic Insights from Molecular Simulations.Biophys J. 2018 Jun 5;114(11):2584-2594. doi: 10.1016/j.bpj.2018.04.023. Biophys J. 2018. PMID: 29874609 Free PMC article.
-
Tracking Membrane Protein Dynamics in Real Time.J Membr Biol. 2021 Feb;254(1):51-64. doi: 10.1007/s00232-020-00165-8. Epub 2021 Jan 7. J Membr Biol. 2021. PMID: 33409541 Free PMC article. Review.
References
-
- Schwab A, Fabian A, Hanley PJ, Stock C. Role of ion channels and transporters in cell migration. Physiol Rev. 2012;92:1865–1913. - PubMed
-
- Finazzi G, Petroutsos D, Tomizioli M, Flori S, Sautron E, Villanova V, Rolland N, Seigneurin-Berny D. Ions channels/transporters and chloroplast regulation. Cell Calcium. 2015;58:86–97. - PubMed
-
- Neher E, Sakmann B. Single-channel currents recorded from membrane of denervated frog muscle fibres. Nature. 1976;260:799–802. - PubMed
-
- Bill RM, Henderson PJ, Iwata S, Kunji ER, Michel H, Neutze R, Newstead S, Poolman B, Tate CG, Vogel H. Overcoming barriers to membrane protein structure determination. Nat Biotechnol. 2011;29:335–340. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
