

A Specialized Peptidoglycan Synthase Promotes Salmonella Cell Division inside Host Cells
- PMID: 29259085
- PMCID: PMC5736910
- DOI: 10.1128/mBio.01685-17
A Specialized Peptidoglycan Synthase Promotes Salmonella Cell Division inside Host Cells
Abstract
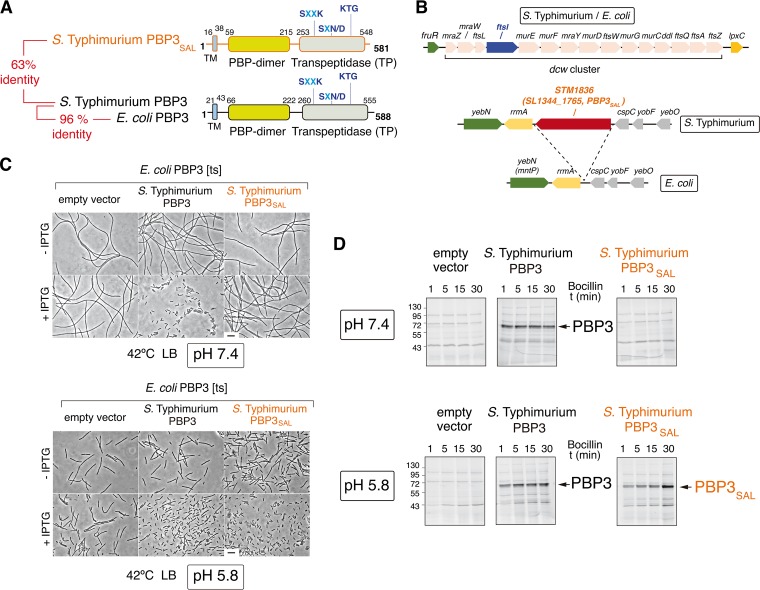
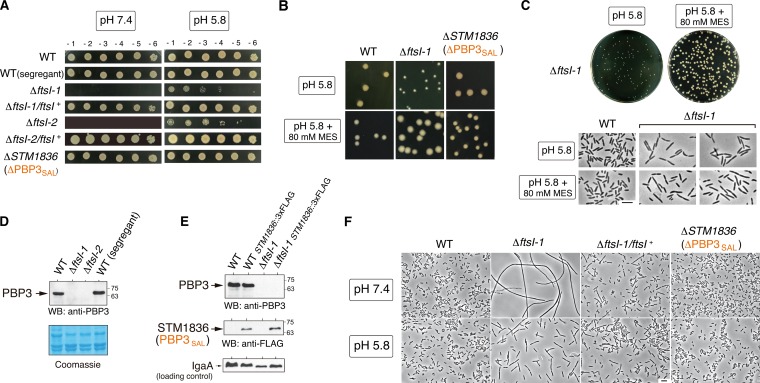
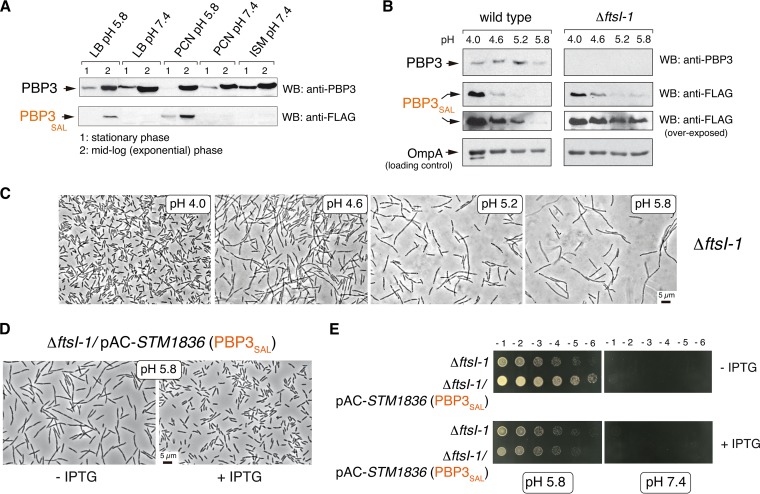
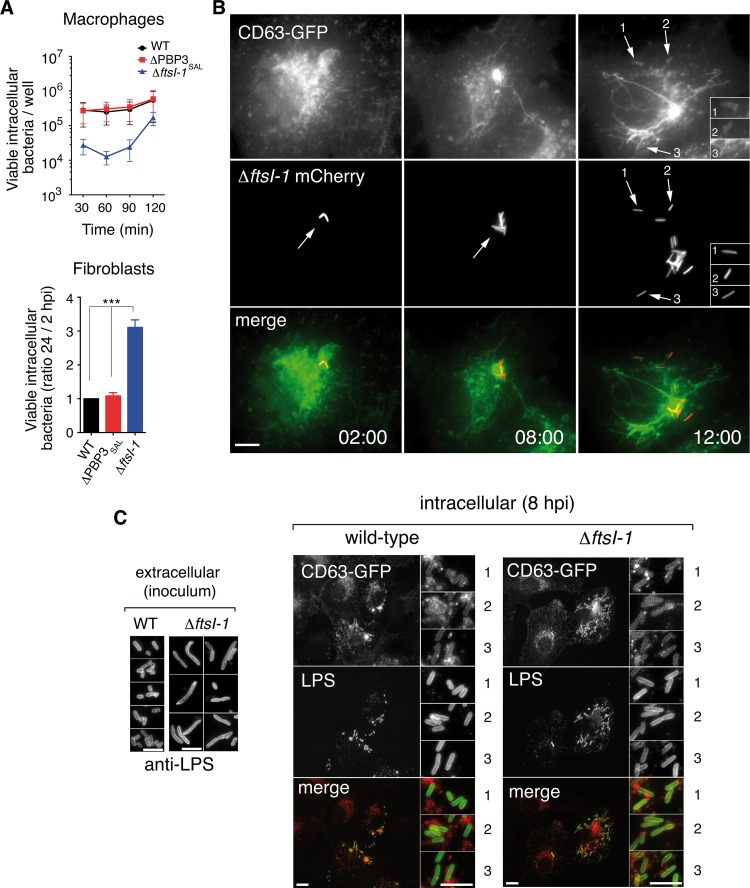
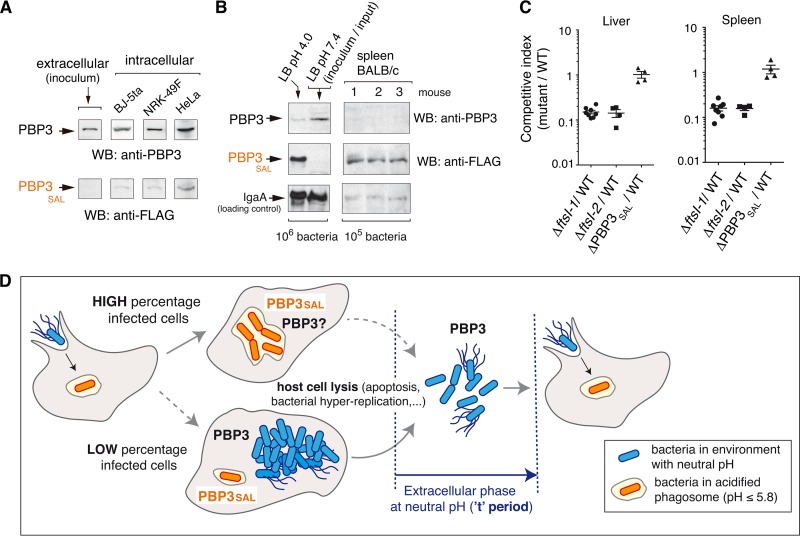
Bacterial cell division has been studied extensively under laboratory conditions. Despite being a key event in the bacterial cell cycle, cell division has not been explored in vivo in bacterial pathogens interacting with their hosts. We discovered in Salmonella enterica serovar Typhimurium a gene absent in nonpathogenic bacteria and encoding a peptidoglycan synthase with 63% identity to penicillin-binding protein 3 (PBP3). PBP3 is an essential cell division-specific peptidoglycan synthase that builds the septum required to separate daughter cells. Since S Typhimurium carries genes that encode a PBP3 paralog-which we named PBP3SAL-and PBP3, we hypothesized that there are different cell division events in host and nonhost environments. To test this, we generated S Typhimurium isogenic mutants lacking PBP3SAL or the hitherto considered essential PBP3. While PBP3 alone promotes cell division under all conditions tested, the mutant producing only PBP3SAL proliferates under acidic conditions (pH ≤ 5.8) but does not divide at neutral pH. PBP3SAL production is tightly regulated with increased levels as bacteria grow in media acidified up to pH 4.0 and in intracellular bacteria infecting eukaryotic cells. PBP3SAL activity is also strictly dependent on acidic pH, as shown by beta-lactam antibiotic binding assays. Live-cell imaging microscopy revealed that PBP3SAL alone is sufficient for S Typhimurium to divide within phagosomes of the eukaryotic cell. Additionally, we detected much larger amounts of PBP3SAL than those of PBP3 in vivo in bacteria colonizing mouse target organs. Therefore, PBP3SAL evolved in S Typhimurium as a specialized peptidoglycan synthase promoting cell division in the acidic intraphagosomal environment.IMPORTANCE During bacterial cell division, daughter cells separate by a transversal structure known as the division septum. The septum is a continuum of the cell wall and therefore is composed of membrane(s) and a peptidoglycan layer. To date, actively growing bacteria were reported to have only a "cell division-specific" peptidoglycan synthase required for the last steps of septum formation and consequently, essential for bacterial life. Here, we discovered that Salmonella enterica has two peptidoglycan synthases capable of synthesizing the division septum. One of these enzymes, PBP3SAL, is present only in bacterial pathogens and evolved in Salmonella to function exclusively in acidic environments. PBP3SAL is used preferentially by Salmonella to promote cell division in vivo in mouse target organs and inside acidified phagosomes. Our data challenge the concept of only one essential cell division-specific peptidoglycan synthase and demonstrate that pathogens can divide in defined host locations using alternative mechanisms.
Keywords: Salmonella; cell division; intracellular pathogens; penicillin-binding proteins; peptidoglycan.
Copyright © 2017 Castanheira et al.
Figures
Comment in
-
Two distinct penicillin binding proteins promote cell division in different Salmonella lifestyles.Microb Cell. 2018 Feb 17;5(3):165-168. doi: 10.15698/mic2018.03.622. Microb Cell. 2018. PMID: 29488513 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
