

A conserved maternal-specific repressive domain in Zelda revealed by Cas9-mediated mutagenesis in Drosophila melanogaster
- PMID: 29261646
- PMCID: PMC5752043
- DOI: 10.1371/journal.pgen.1007120
A conserved maternal-specific repressive domain in Zelda revealed by Cas9-mediated mutagenesis in Drosophila melanogaster
Abstract
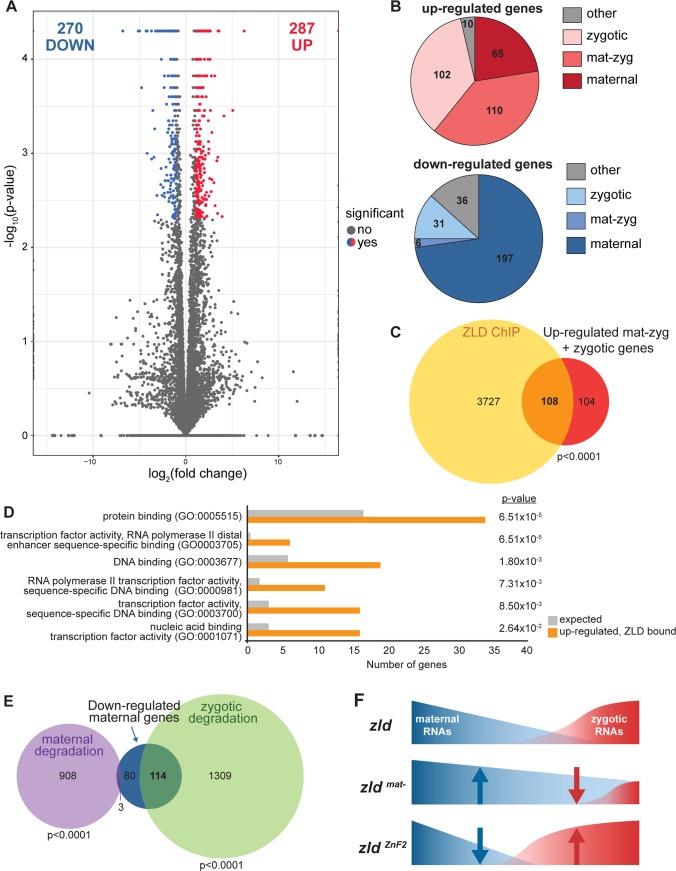
In nearly all metazoans, the earliest stages of development are controlled by maternally deposited mRNAs and proteins. The zygotic genome becomes transcriptionally active hours after fertilization. Transcriptional activation during this maternal-to-zygotic transition (MZT) is tightly coordinated with the degradation of maternally provided mRNAs. In Drosophila melanogaster, the transcription factor Zelda plays an essential role in widespread activation of the zygotic genome. While Zelda expression is required both maternally and zygotically, the mechanisms by which it functions to remodel the embryonic genome and prepare the embryo for development remain unclear. Using Cas9-mediated genome editing to generate targeted mutations in the endogenous zelda locus, we determined the functional relevance of protein domains conserved amongst Zelda orthologs. We showed that neither a conserved N-terminal zinc finger nor an acidic patch were required for activity. Similarly, a previously identified splice isoform of zelda is dispensable for viability. By contrast, we identified a highly conserved zinc-finger domain that is essential for the maternal, but not zygotic functions of Zelda. Animals homozygous for mutations in this domain survived to adulthood, but embryos inheriting these loss-of-function alleles from their mothers died late in embryogenesis. These mutations did not interfere with the capacity of Zelda to activate transcription in cell culture. Unexpectedly, these mutations generated a hyperactive form of the protein and enhanced Zelda-dependent gene expression. These data have defined a protein domain critical for controlling Zelda activity during the MZT, but dispensable for its roles later in development, for the first time separating the maternal and zygotic requirements for Zelda. This demonstrates that highly regulated levels of Zelda activity are required for establishing the developmental program during the MZT. We propose that tightly regulated gene expression is essential to navigate the MZT and that failure to precisely execute this developmental program leads to embryonic lethality.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Comment in
-
Maternal or zygotic: Unveiling the secrets of the Pancrustacea transcription factor zelda.PLoS Genet. 2018 Mar 1;14(3):e1007201. doi: 10.1371/journal.pgen.1007201. eCollection 2018 Mar. PLoS Genet. 2018. PMID: 29494591 Free PMC article. No abstract available.
Similar articles
-
Transcriptional activation is a conserved feature of the early embryonic factor Zelda that requires a cluster of four zinc fingers for DNA binding and a low-complexity activation domain.J Biol Chem. 2015 Feb 6;290(6):3508-18. doi: 10.1074/jbc.M114.602292. Epub 2014 Dec 23. J Biol Chem. 2015. PMID: 25538246 Free PMC article.
-
Zelda binding in the early Drosophila melanogaster embryo marks regions subsequently activated at the maternal-to-zygotic transition.PLoS Genet. 2011 Oct;7(10):e1002266. doi: 10.1371/journal.pgen.1002266. Epub 2011 Oct 20. PLoS Genet. 2011. PMID: 22028662 Free PMC article.
-
Zelda and the maternal-to-zygotic transition in cockroaches.FEBS J. 2019 Aug;286(16):3206-3221. doi: 10.1111/febs.14856. Epub 2019 Apr 30. FEBS J. 2019. PMID: 30993896
-
Regulatory principles governing the maternal-to-zygotic transition: insights from Drosophila melanogaster.Open Biol. 2018 Dec;8(12):180183. doi: 10.1098/rsob.180183. Open Biol. 2018. PMID: 30977698 Free PMC article. Review.
-
Noninheritable Maternal Factors Useful for Genetic Manipulation in Mammals.Results Probl Cell Differ. 2017;63:495-510. doi: 10.1007/978-3-319-60855-6_21. Results Probl Cell Differ. 2017. PMID: 28779331 Review.
Cited by
-
Stepwise modifications of transcriptional hubs link pioneer factor activity to a burst of transcription.Nat Commun. 2023 Aug 10;14(1):4848. doi: 10.1038/s41467-023-40485-6. Nat Commun. 2023. PMID: 37563108 Free PMC article.
-
Comparative transcriptome analyses of the Drosophila pupal eye.G3 (Bethesda). 2021 Jan 18;11(1):jkaa003. doi: 10.1093/g3journal/jkaa003. G3 (Bethesda). 2021. PMID: 33561221 Free PMC article.
-
The pioneer transcription factor Zelda controls the exit from regeneration and restoration of patterning in Drosophila.Sci Adv. 2025 Jun 6;11(23):eads5743. doi: 10.1126/sciadv.ads5743. Epub 2025 Jun 6. Sci Adv. 2025. PMID: 40479065 Free PMC article.
-
Maternal or zygotic: Unveiling the secrets of the Pancrustacea transcription factor zelda.PLoS Genet. 2018 Mar 1;14(3):e1007201. doi: 10.1371/journal.pgen.1007201. eCollection 2018 Mar. PLoS Genet. 2018. PMID: 29494591 Free PMC article. No abstract available.
-
CLAMP and Zelda function together to promote Drosophila zygotic genome activation.Elife. 2021 Aug 3;10:e69937. doi: 10.7554/eLife.69937. Elife. 2021. PMID: 34342574 Free PMC article.
References
-
- Schier AF. The maternal-zygotic transition: death and birth of RNAs. Science. 2007;316(5823):406–7. doi: 10.1126/science.1140693 - DOI - PubMed
-
- Tadros W, Lipshitz HD. The maternal-to-zygotic transition: a play in two acts. Development. 2009;136(18):3033–42. doi: 10.1242/dev.033183 - DOI - PubMed
-
- Harrison MM, Eisen MB. Transcriptional Activation of the Zygotic Genome in Drosophila. Matern Zygotic Transit. 2015;113:85–112. - PubMed
-
- Lee MT, Bonneau AR, Giraldez AJ. Zygotic genome activation during the maternal-to-zygotic transition. Annu Rev Cell Dev Biol. 2014;30:581–613. doi: 10.1146/annurev-cellbio-100913-013027 - DOI - PMC - PubMed
-
- Bashirullah A, Halsell SR, Cooperstock RL, Kloc M, Karaiskakis A, Fisher WW, et al. Joint action of two RNA degradation pathways controls the timing of maternal transcript elimination at the midblastula transition in Drosophila melanogaster. EMBO J. 1999. May 4;18(9):2610–20. doi: 10.1093/emboj/18.9.2610 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
