

Association with Aurora-A Controls N-MYC-Dependent Promoter Escape and Pause Release of RNA Polymerase II during the Cell Cycle
- PMID: 29262328
- PMCID: PMC5746598
- DOI: 10.1016/j.celrep.2017.11.090
Association with Aurora-A Controls N-MYC-Dependent Promoter Escape and Pause Release of RNA Polymerase II during the Cell Cycle
Abstract
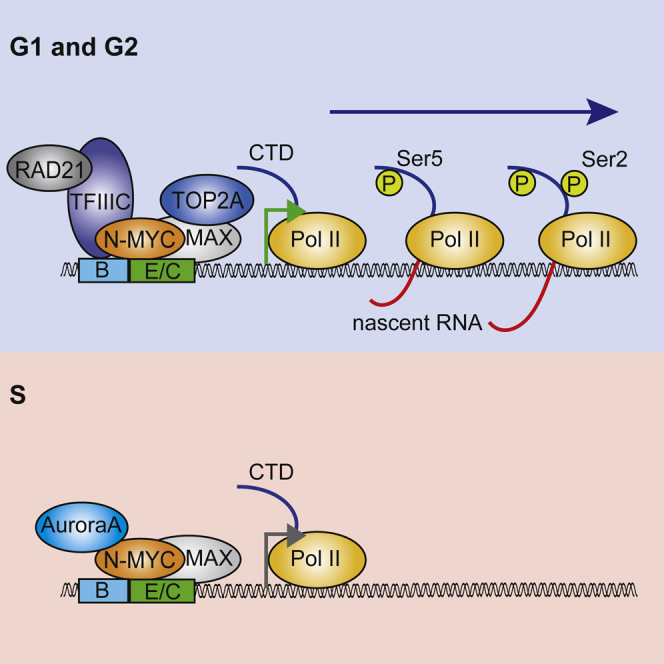
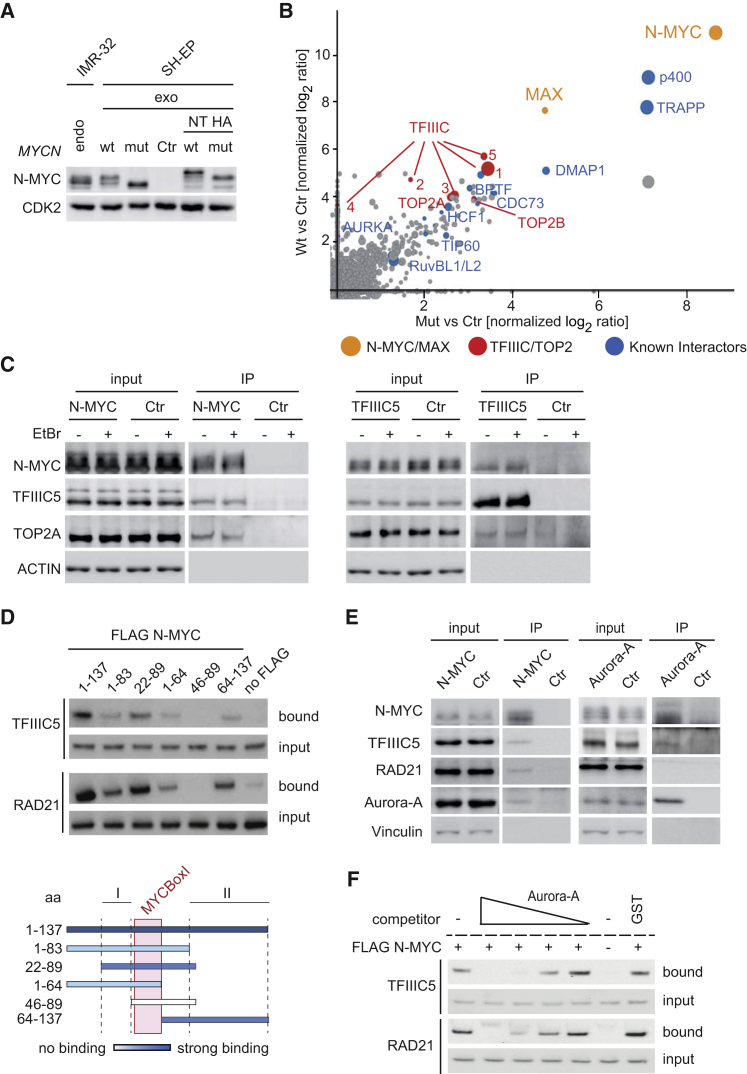
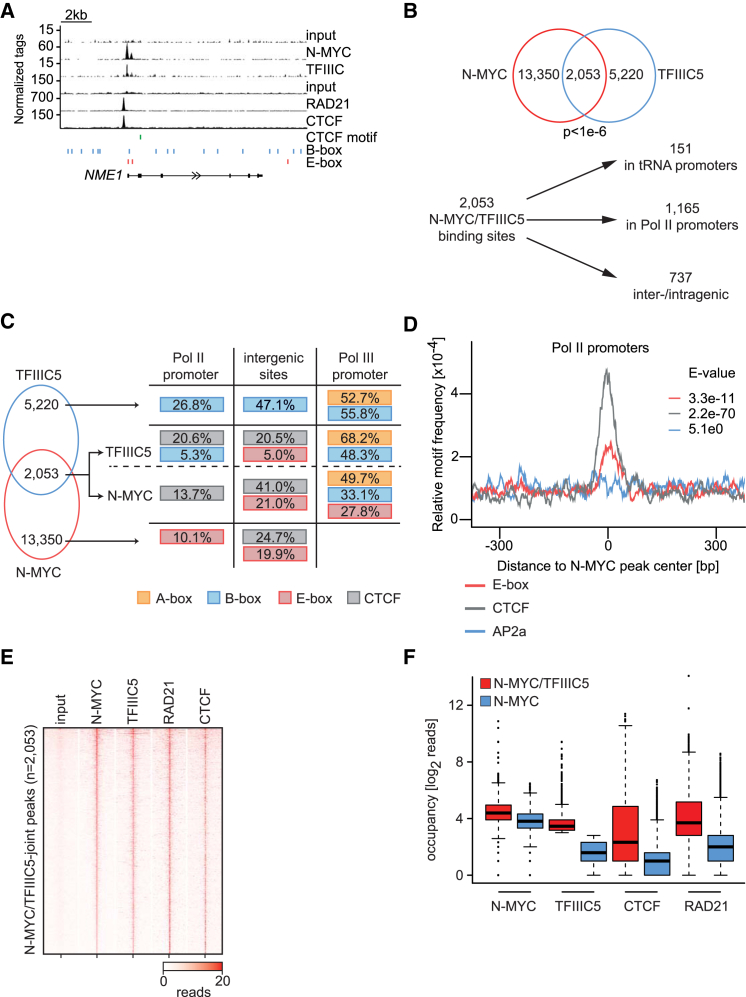
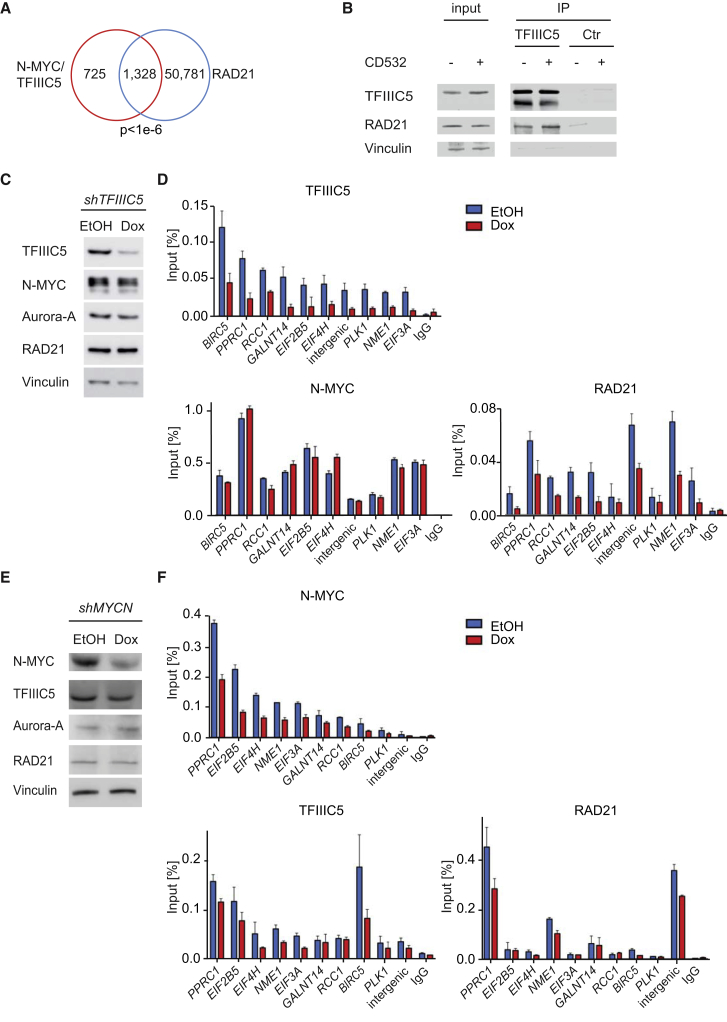
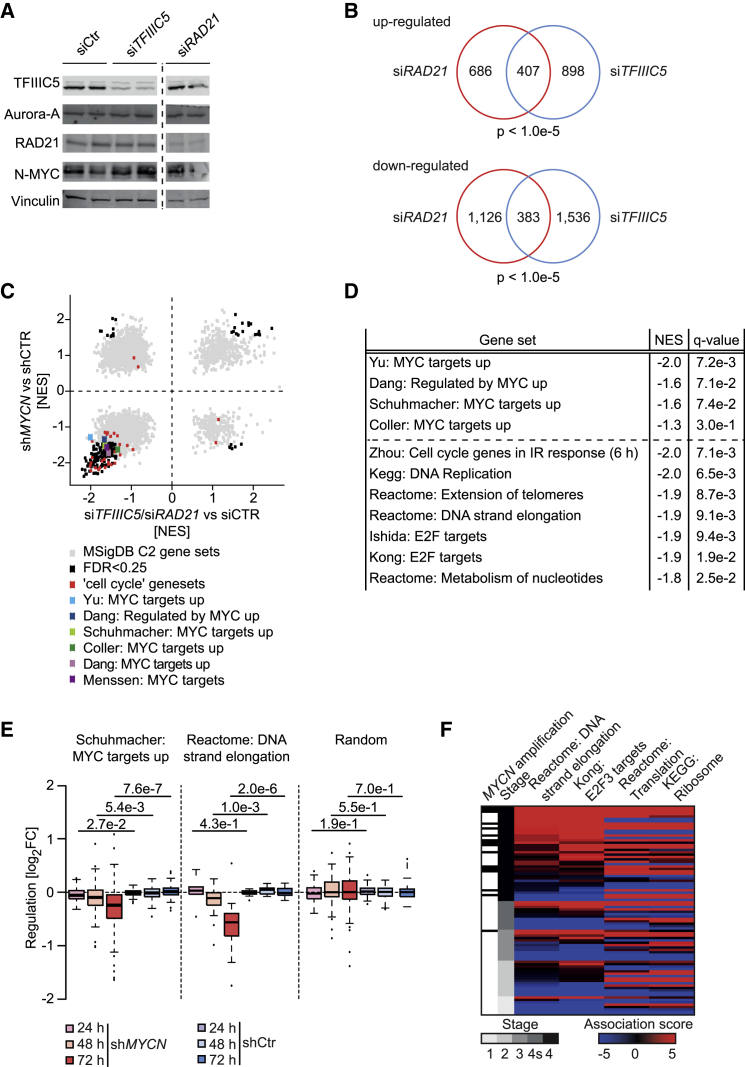
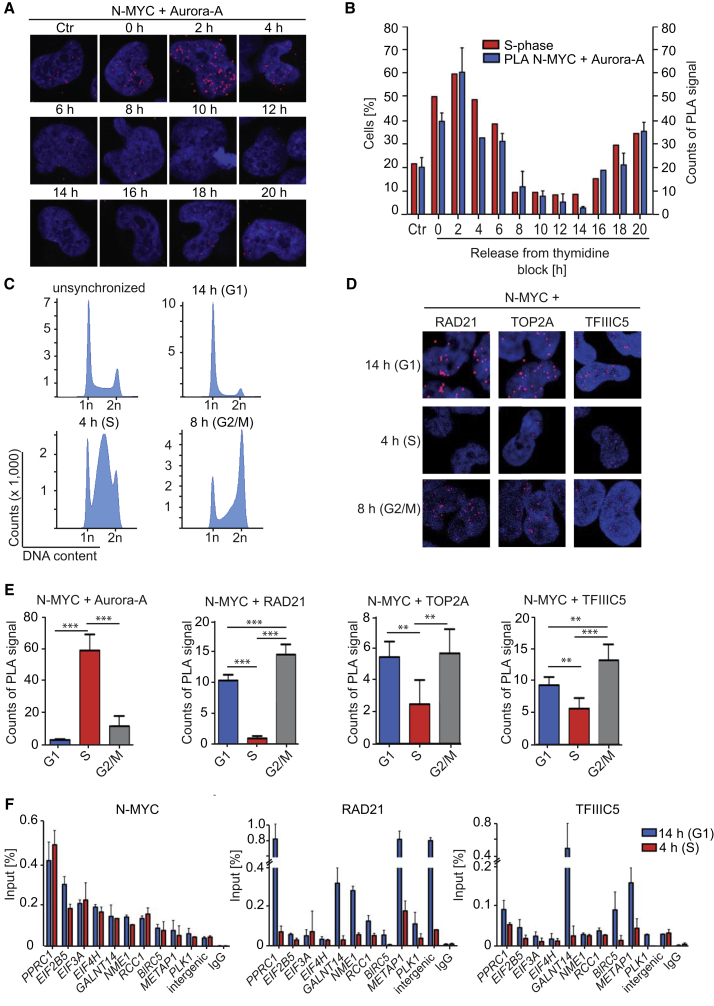
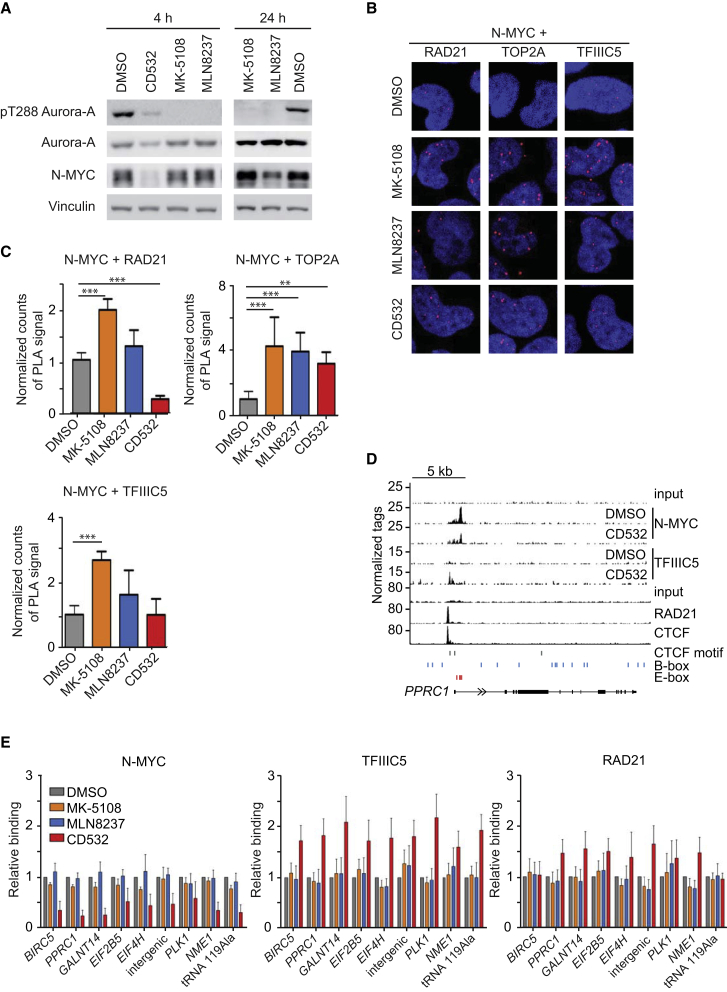
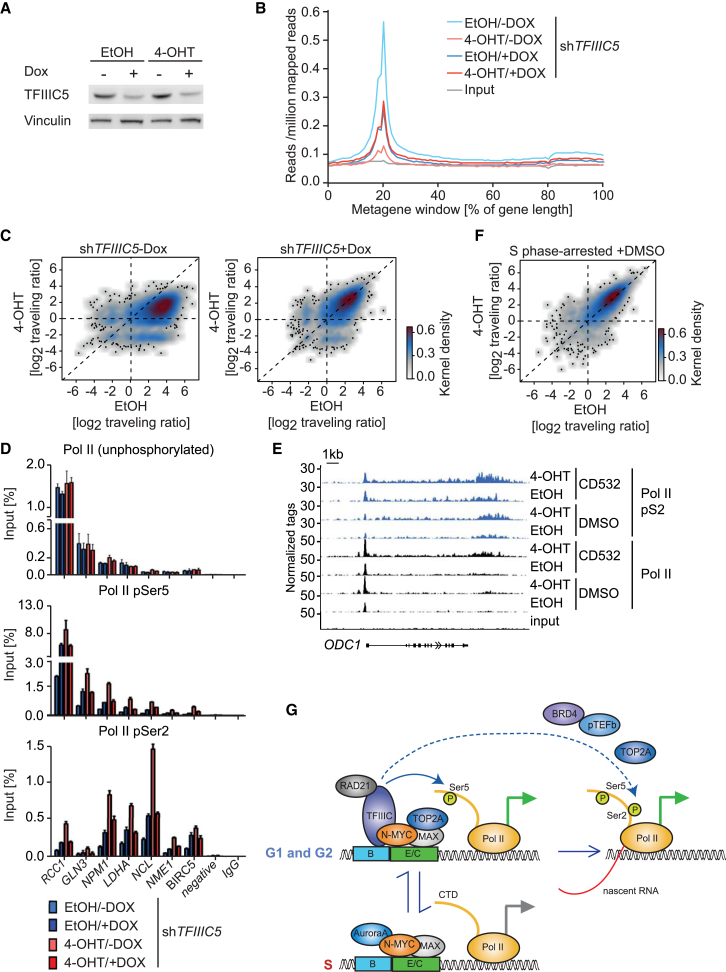
MYC proteins bind globally to active promoters and promote transcriptional elongation by RNA polymerase II (Pol II). To identify effector proteins that mediate this function, we performed mass spectrometry on N-MYC complexes in neuroblastoma cells. The analysis shows that N-MYC forms complexes with TFIIIC, TOP2A, and RAD21, a subunit of cohesin. N-MYC and TFIIIC bind to overlapping sites in thousands of Pol II promoters and intergenic regions. TFIIIC promotes association of RAD21 with N-MYC target sites and is required for N-MYC-dependent promoter escape and pause release of Pol II. Aurora-A competes with binding of TFIIIC and RAD21 to N-MYC in vitro and antagonizes association of TOP2A, TFIIIC, and RAD21 with N-MYC during S phase, blocking N-MYC-dependent release of Pol II from the promoter. Inhibition of Aurora-A in S phase restores RAD21 and TFIIIC binding to chromatin and partially restores N-MYC-dependent transcriptional elongation. We propose that complex formation with Aurora-A controls N-MYC function during the cell cycle.
Keywords: Aurora-A; MYC; N-MYC; RAD21; TFIIIC; neuroblastoma; pause release.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Beltran H. The N-MYC oncogene: maximizing its targets, regulation, and therapeutic potential. Mol. Cancer Res. 2014;12:815–822. - PubMed
-
- Carrière L., Graziani S., Alibert O., Ghavi-Helm Y., Boussouar F., Humbertclaude H., Jounier S., Aude J.C., Keime C., Murvai J. Genomic binding of Pol III transcription machinery and relationship with TFIIS transcription factor distribution in mouse embryonic stem cells. Nucleic Acids Res. 2012;40:270–283. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
