

Human papillomavirus type 18 E5 oncogene supports cell cycle progression and impairs epithelial differentiation by modulating growth factor receptor signalling during the virus life cycle
- PMID: 29262586
- PMCID: PMC5732752
- DOI: 10.18632/oncotarget.21658
Human papillomavirus type 18 E5 oncogene supports cell cycle progression and impairs epithelial differentiation by modulating growth factor receptor signalling during the virus life cycle
Abstract
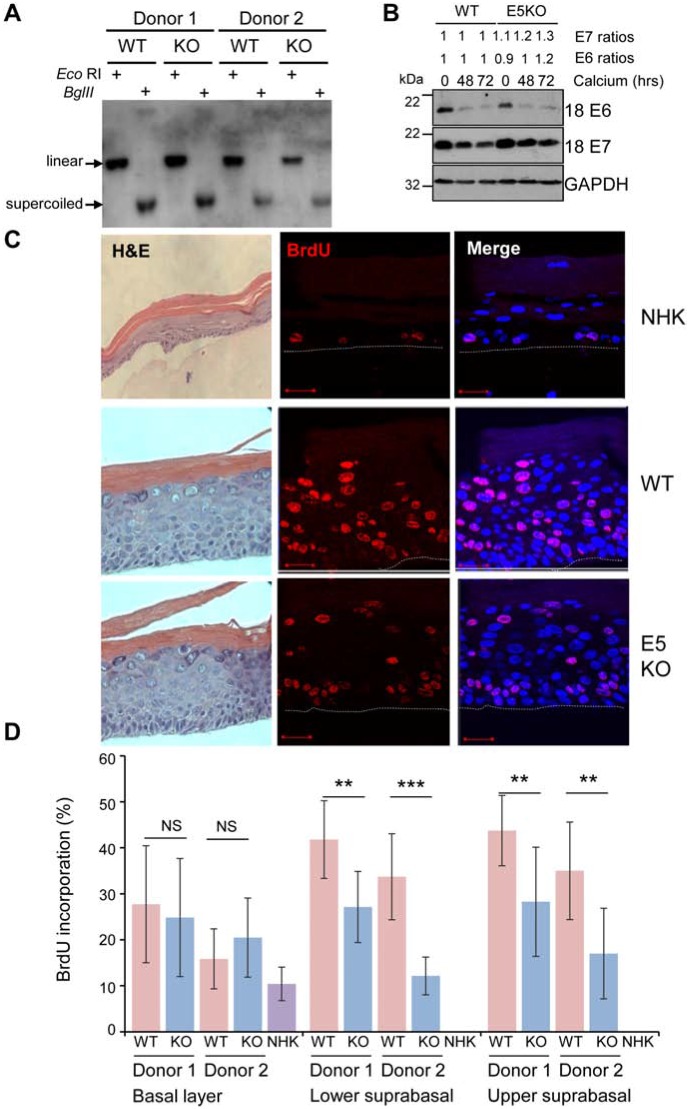
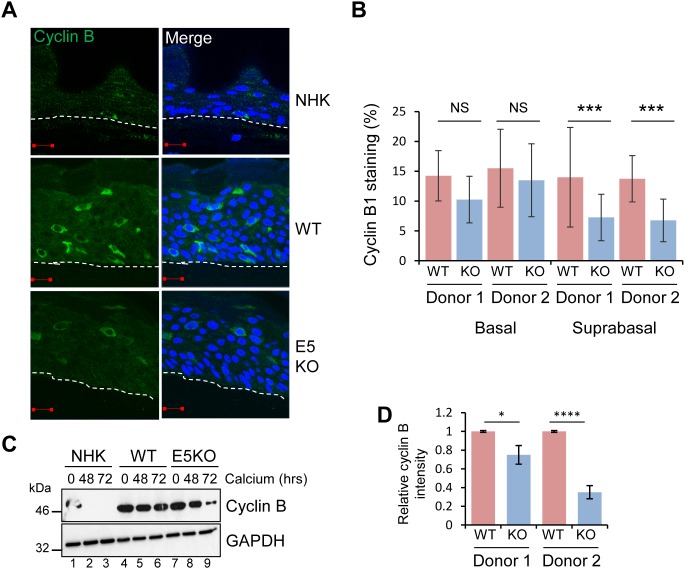
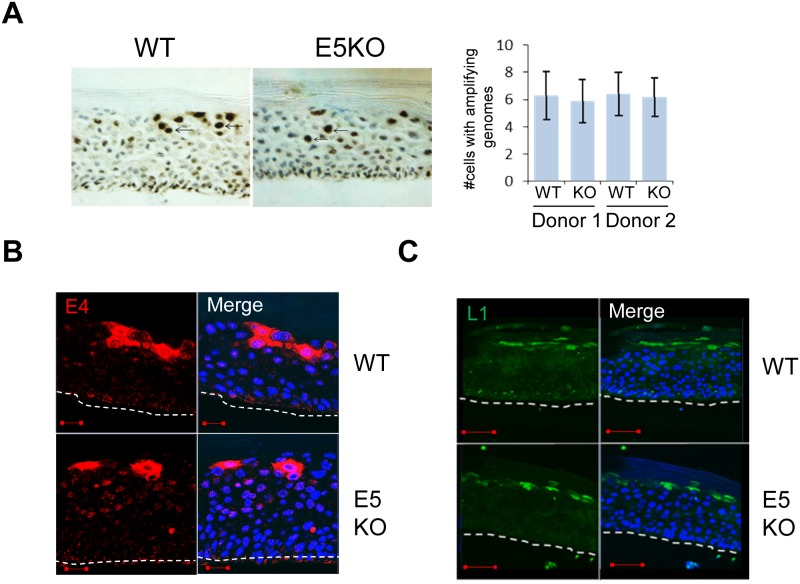
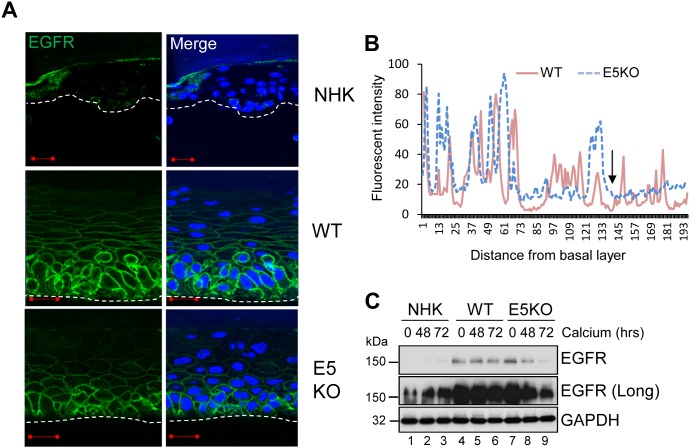
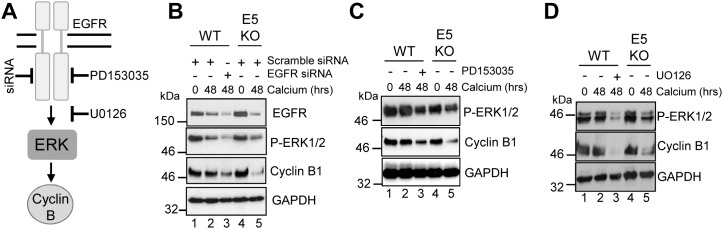
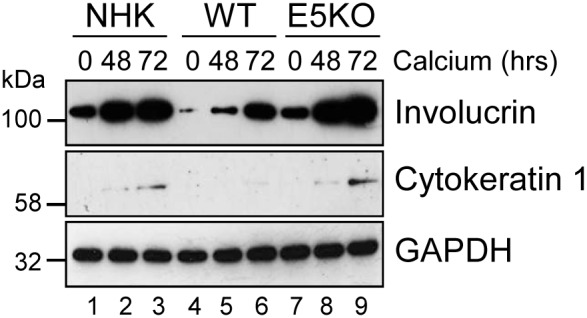
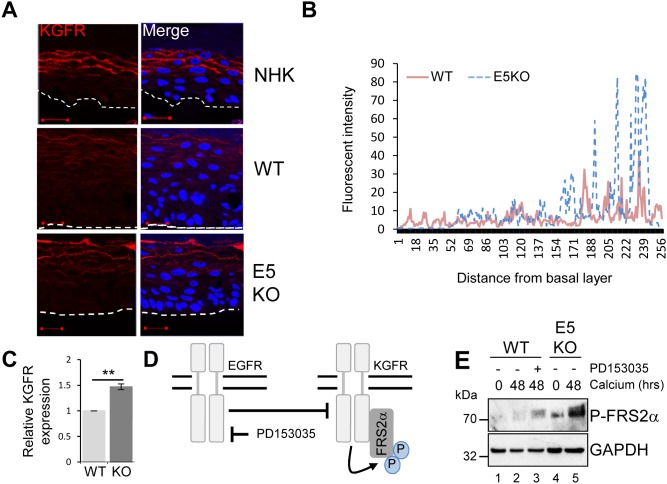
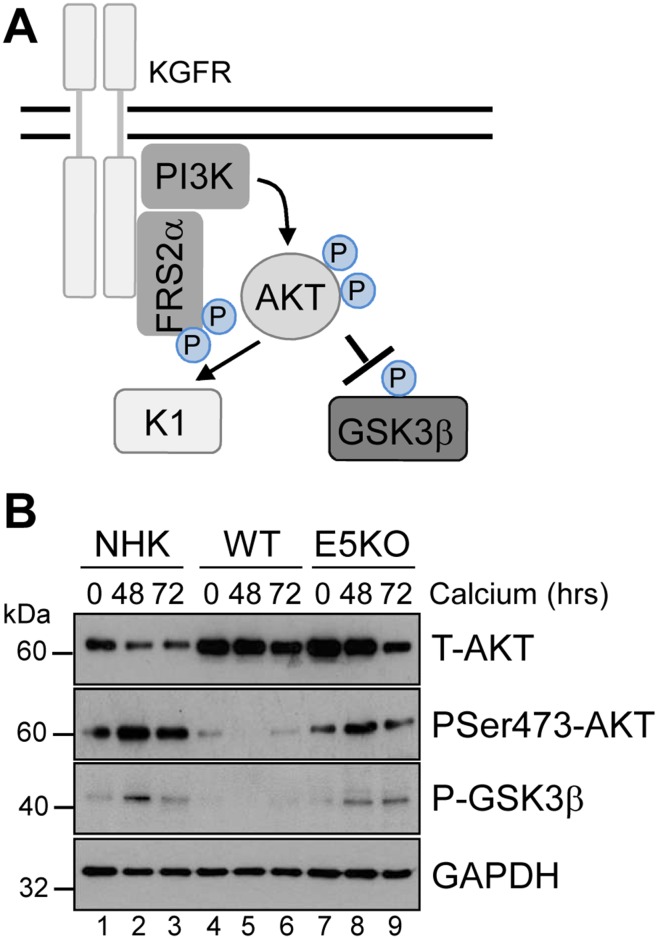
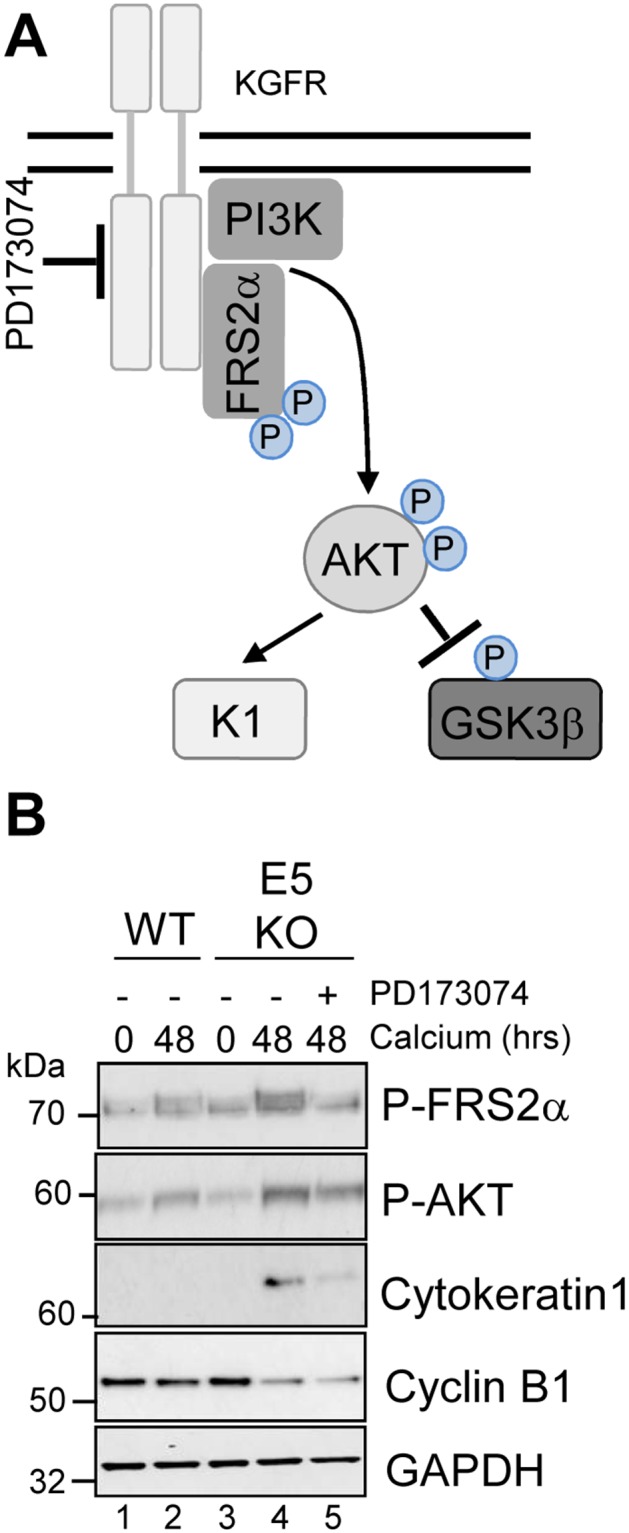
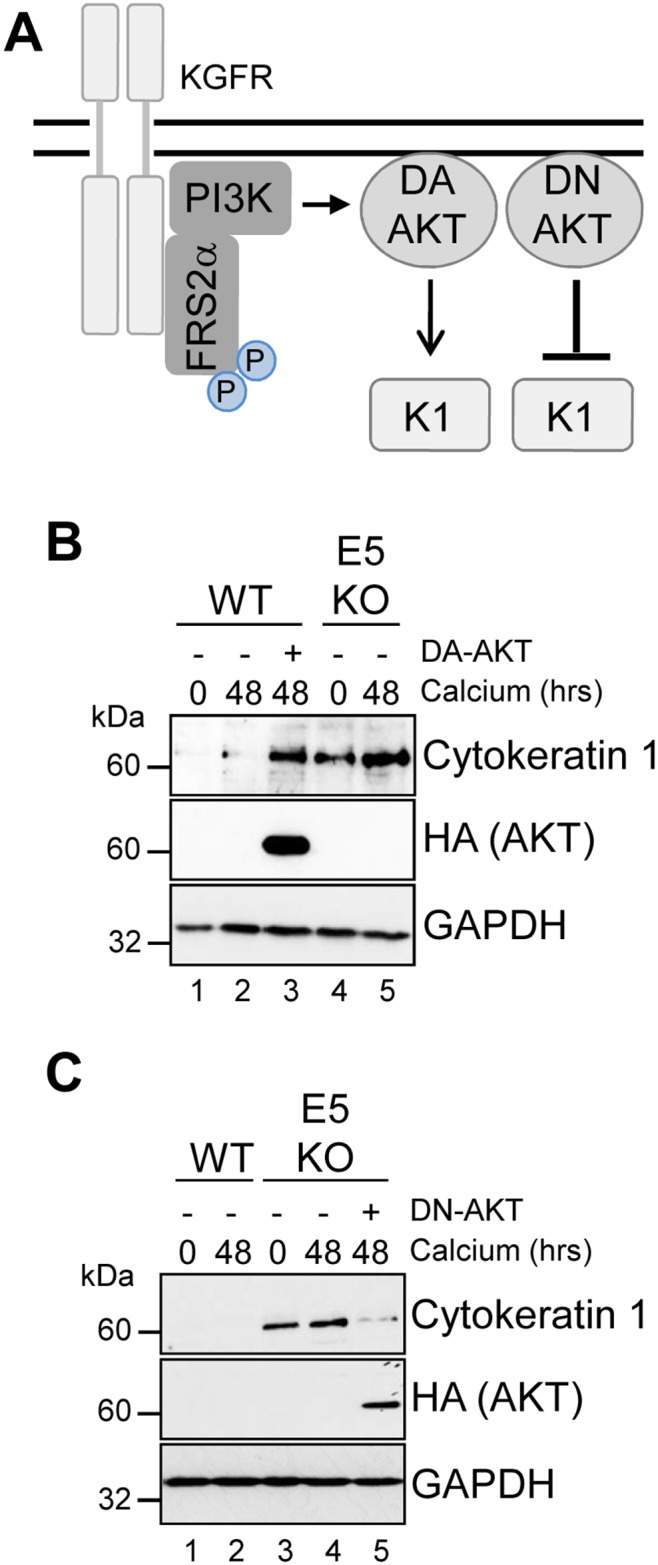
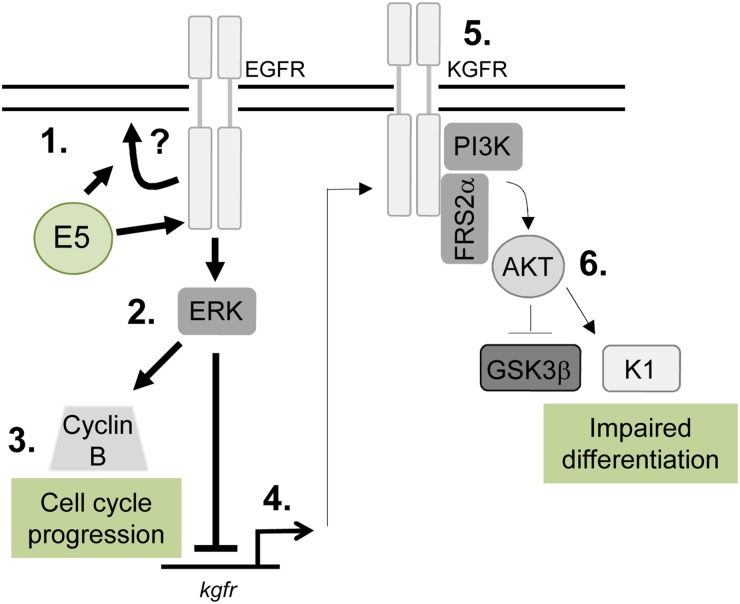
Deregulation of proliferation and differentiation-dependent signalling pathways is a hallmark of human papillomavirus (HPV) infection. Although the manipulation of these pathways by E6 and E7 has been extensively studied, controversies surround the role of the E5 oncoprotein during a productive virus life cycle. By integrating primary keratinocytes harbouring wild type or E5 knockout HPV18 genomes with pharmacological and gain/loss of function models, this study aimed to provide molecular information about the role of E5 in epithelial proliferation and differentiation. We show that E5 contributes to cell cycle progression and unscheduled host DNA synthesis in differentiating keratinocytes. E5 function correlates with increased EGFR activation in differentiating cells and blockade of this pathway impairs differentiation-dependent cell cycle progression of HPV18 containing cells. Our findings provide a functional requirement of enhanced EGFR signalling for suprabasal cellular DNA synthesis during the virus life cycle. They also reveal an unrecognised contribution of E5 towards the impaired keratinocyte differentiation observed during a productive HPV infection. E5 suppresses a signalling axis consisting of the keratinocyte growth factor receptor (KGFR) pathway. Inhibition of this pathway compensates for the loss of E5 in knockout cells and re-instates the delay in differentiation. The negative regulation of KGFR involves suppression by the EGFR pathway. Thus our data reveal an unappreciated role for E5-mediated EGFR signalling in orchestrating the balance between proliferation and differentiation in suprabasal cells.
Keywords: E5; EGFR; HPV; differentiation; proliferation.
Conflict of interest statement
CONFLICTS OF INTEREST The authors declare that there is no conflict of interest.
Figures
References
-
- Doorbar J, Quint W, Banks L, Bravo IG, Stoler M, Broker TR, Stanley MA. The biology and life-cycle of human papillomaviruses. Vaccine. 2012;30:F55–F70. http://doi.org/10.1016/j.vaccine.2012.06.083. - DOI - PubMed
-
- Roberts S. Papillomaviruses. eLS. Chichester, UK: John Wiley & Sons, Ltd; 2001. pp. 1–11. http://doi.org/10.1002/9780470015902.a0000422.pub3. - DOI
-
- Clifford GM, Smith JS, Aguado T, Franceschi S. Comparison of HPV type distribution in high-grade cervical lesions and cervical cancer: a meta-analysis. Br J Cancer. 2003;89:101–105. http://doi.org/10.1038/sj.bjc.6601024. - DOI - PMC - PubMed
-
- de Sanjose S, Quint WG, Alemany L, Geraets DT, Klaustermeier JE, Lloveras B, Tous S, Felix A, Bravo LE, Shin HR, Vallejos CS, de Ruiz PA, Lima MA, et al. Human papillomavirus genotype attribution in invasive cervical cancer: a retrospective cross-sectional worldwide study. Lancet Oncol. 2010;11:1048–1056. http://doi.org/10.1016/S1470-2045(10)70230-8. - DOI - PubMed
-
- Dumesic PA, Scholl FA, Barragan DI, Khavari PA. Erk1/2 MAP kinases are required for epidermal G2/M progression. J Cell Biol. 2009;185:409–422. http://doi.org/10.1083/jcb.200804038. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
