

MUC1-C Induces PD-L1 and Immune Evasion in Triple-Negative Breast Cancer
- PMID: 29263152
- PMCID: PMC5754244
- DOI: 10.1158/0008-5472.CAN-17-1636
MUC1-C Induces PD-L1 and Immune Evasion in Triple-Negative Breast Cancer
Abstract
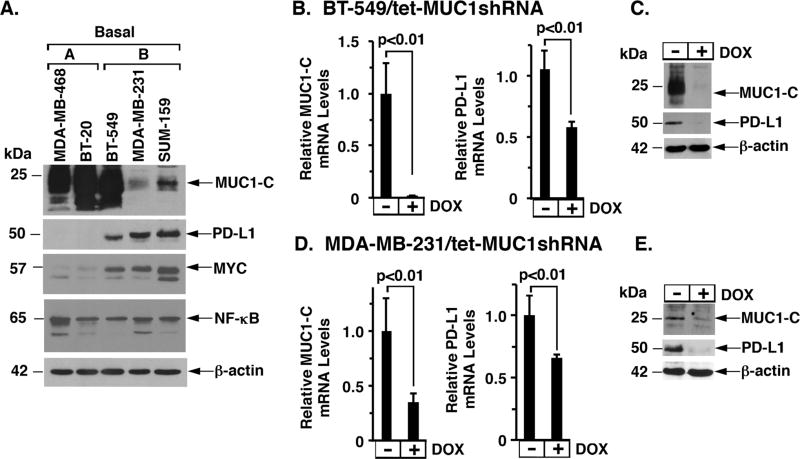
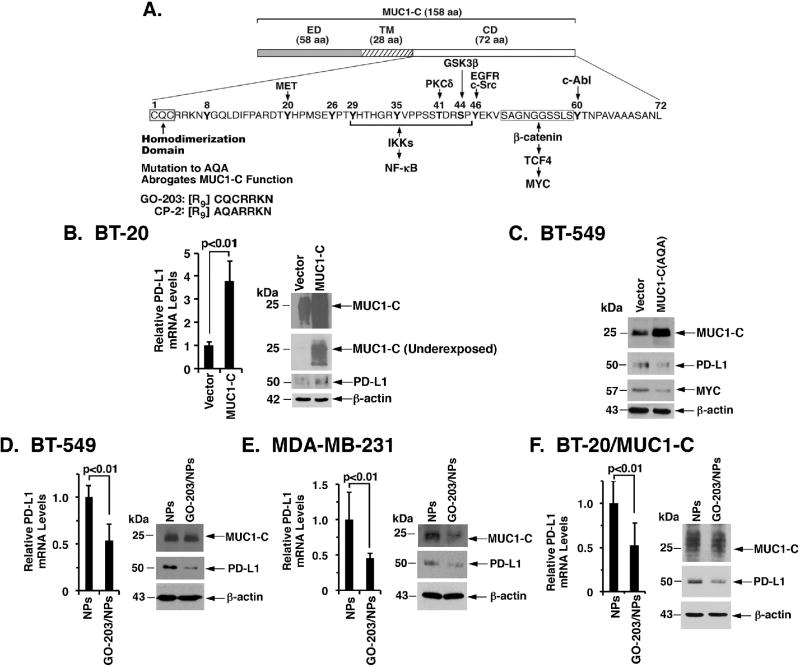
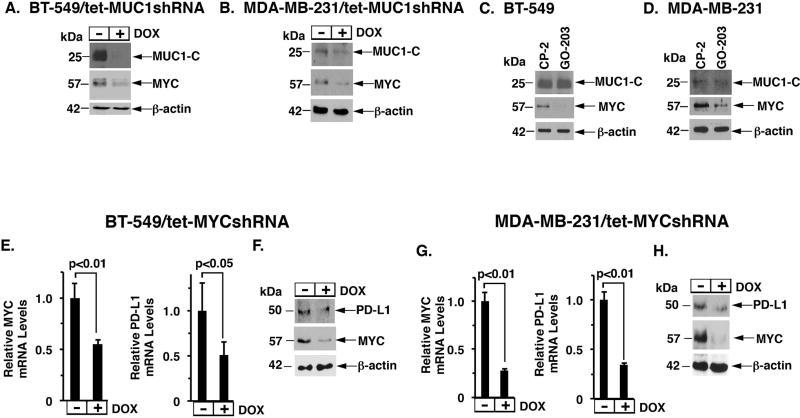
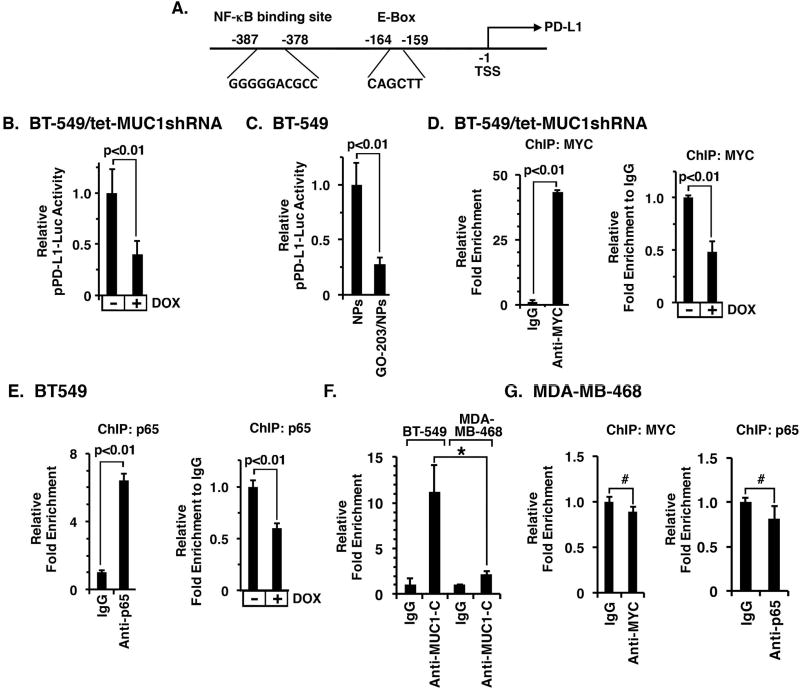
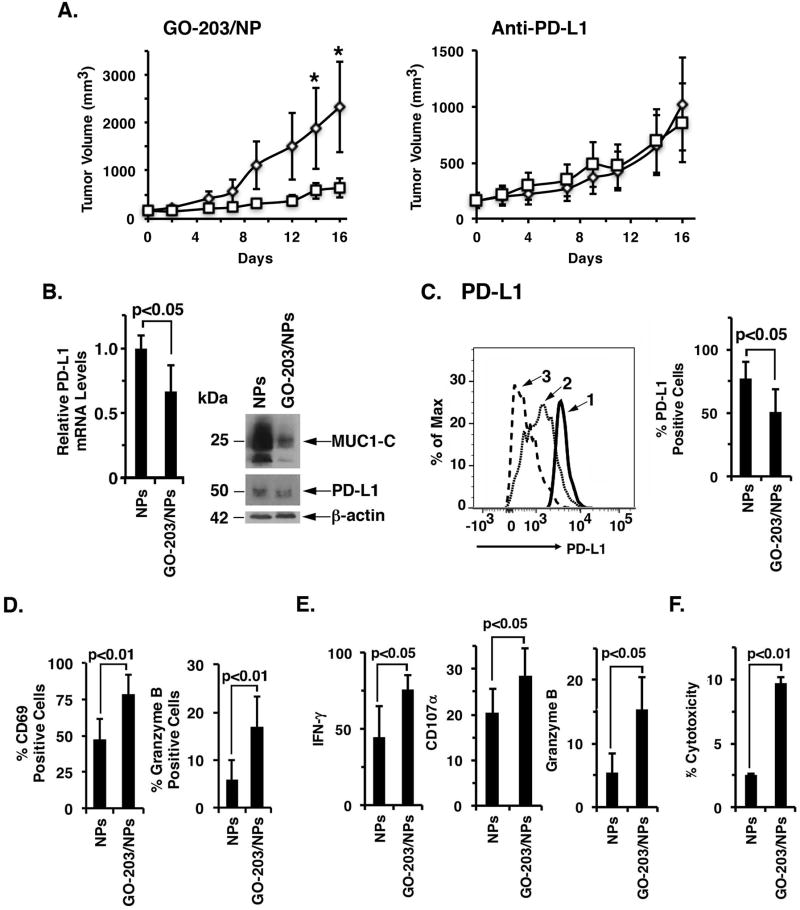
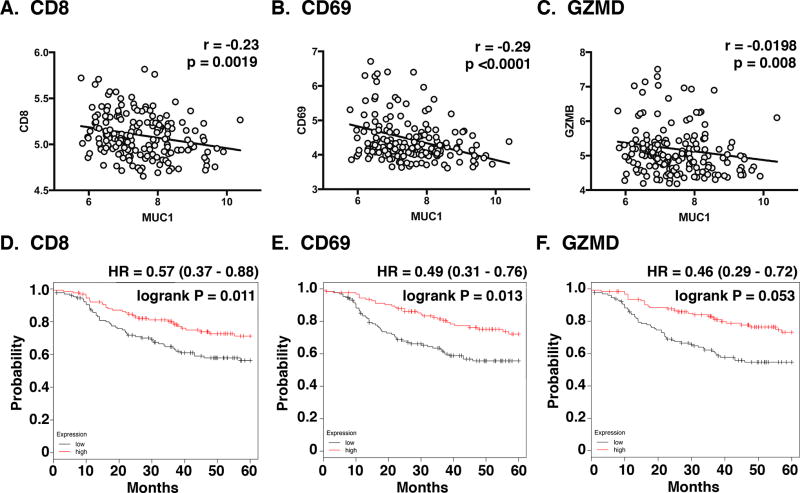
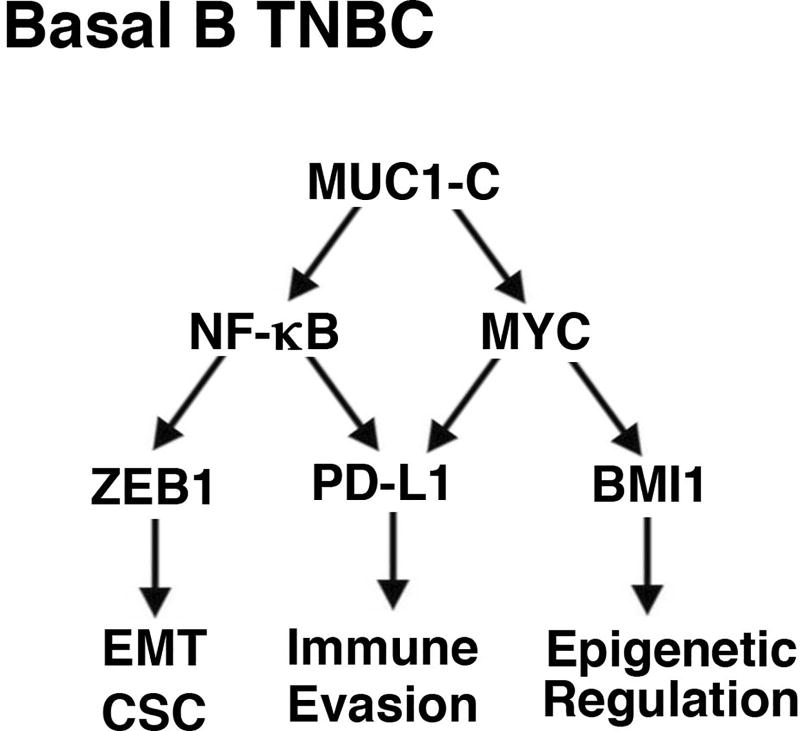
The immune checkpoint ligand PD-L1 and the transmembrane mucin MUC1 are upregulated in triple-negative breast cancer (TNBC), where they contribute to its aggressive pathogenesis. Here, we report that genetic or pharmacological targeting of the oncogenic MUC1 subunit MUC1-C is sufficient to suppress PD-L1 expression in TNBC cells. Mechanistic investigations showed that MUC1-C acted to elevate PD-L1 transcription by recruitment of MYC and NF-κB p65 to the PD-L1 promoter. In an immunocompetent model of TNBC in which Eo771/MUC1-C cells were engrafted into MUC1 transgenic mice, we showed that targeting MUC1-C associated with PD-L1 suppression, increases in tumor-infiltrating CD8+ T cells and tumor cell killing. MUC1 expression in TNBCs also correlated inversely with CD8, CD69, and GZMB, and downregulation of these markers associated with decreased survival. Taken together, our findings show how MUC1 contributes to immune escape in TNBC, and they offer a rationale to target MUC1-C as a novel immunotherapeutic approach for TNBC treatment.Significance: These findings show how upregulation of the transmembrane mucin MUC1 contributes to immune escape in an aggressive form of breast cancer, with potential implications for a novel immunotherapeutic approach. Cancer Res; 78(1); 205-15. ©2017 AACR.
©2017 American Association for Cancer Research.
Conflict of interest statement
Figures
References
-
- Beckers RK, Selinger CI, Vilain R, Madore J, Wilmott JS, Harvey K, et al. Programmed death ligand 1 expression in triple-negative breast cancer is associated with tumour-infiltrating lymphocytes and improved outcome. Histopathology. 2016;69:25–34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
