

A human microglia-like cellular model for assessing the effects of neurodegenerative disease gene variants
- PMID: 29263232
- PMCID: PMC5945290
- DOI: 10.1126/scitranslmed.aai7635
A human microglia-like cellular model for assessing the effects of neurodegenerative disease gene variants
Abstract
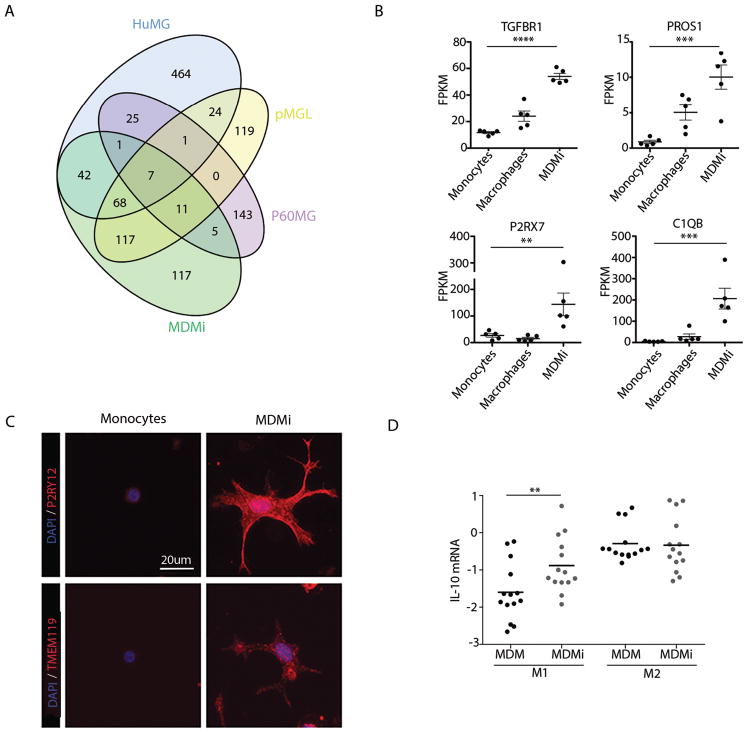
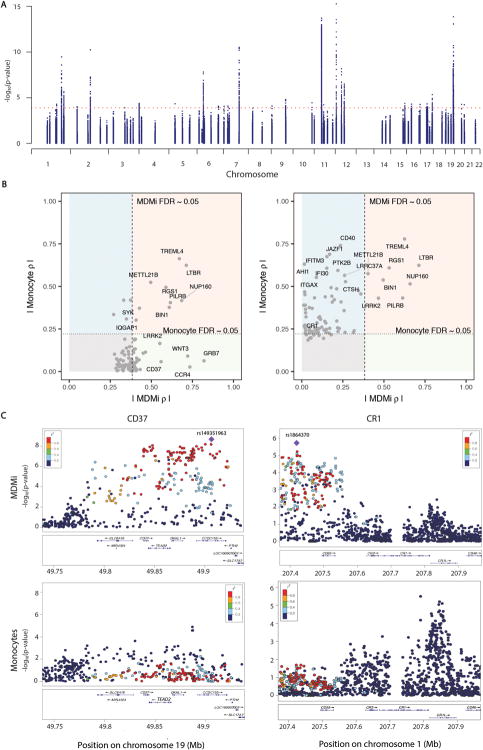
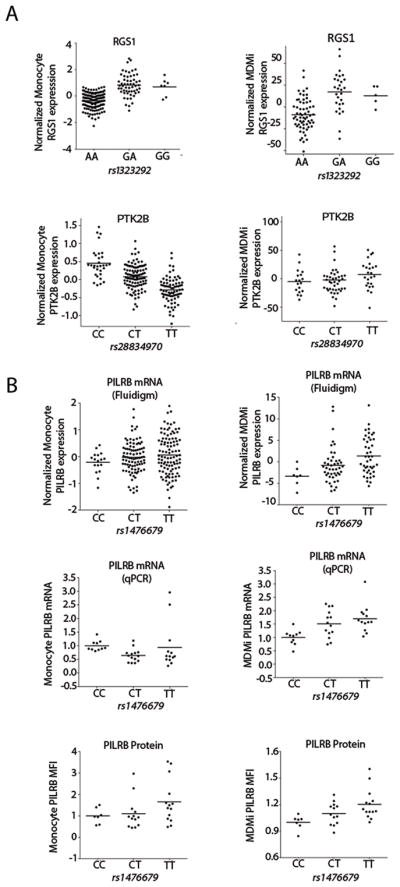
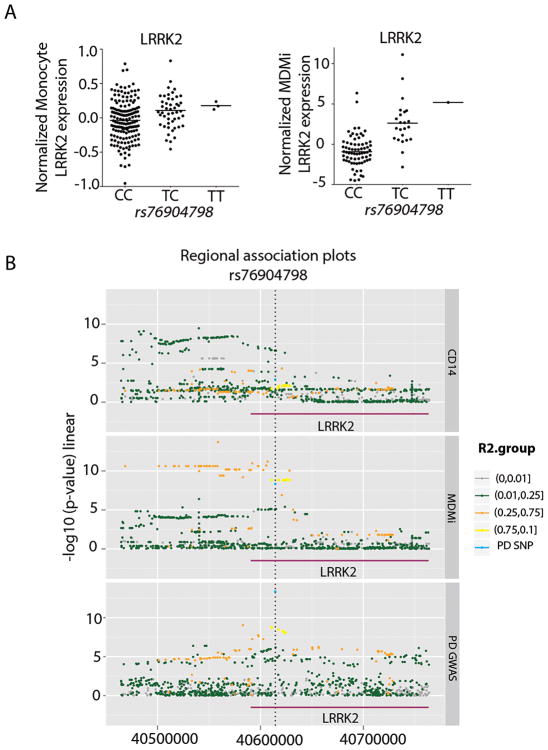
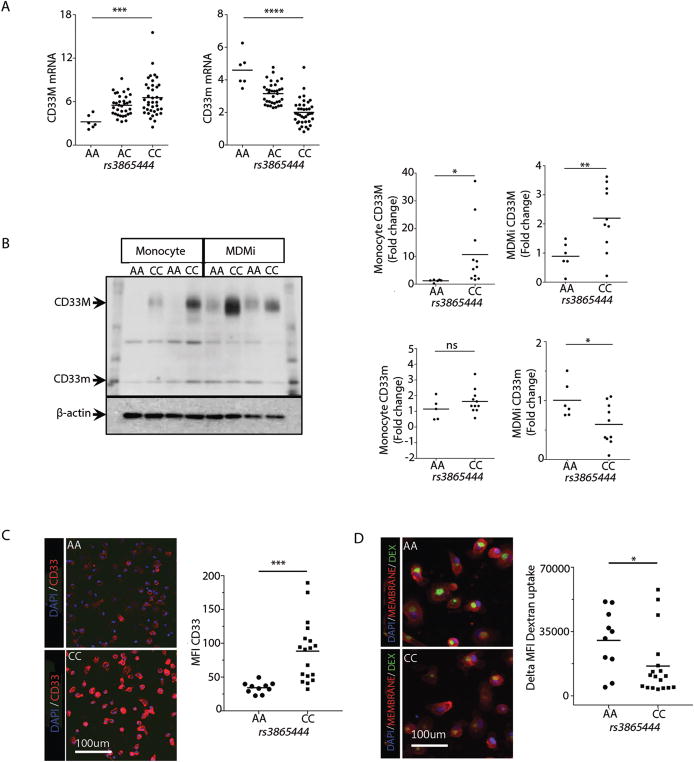
Microglia are emerging as a key cell type in neurodegenerative diseases, yet human microglia are challenging to study in vitro. We developed an in vitro cell model system composed of human monocyte-derived microglia-like (MDMi) cells that recapitulated key aspects of microglia phenotype and function. We then used this model system to perform an expression quantitative trait locus (eQTL) study examining 94 genes from loci associated with Alzheimer's disease, Parkinson's disease, and multiple sclerosis. We found six loci (CD33, PILRB, NUP160, LRRK2, RGS1, and METTL21B) in which the risk haplotype drives the association with both disease susceptibility and altered expression of a nearby gene (cis-eQTL). In the PILRB and LRRK2 loci, the cis-eQTL was found in the MDMi cells but not in human peripheral blood monocytes, suggesting that differentiation of monocytes into microglia-like cells led to the acquisition of a cellular state that could reveal the functional consequences of certain genetic variants. We further validated the effect of risk haplotypes at the protein level for PILRB and CD33, and we confirmed that the CD33 risk haplotype altered phagocytosis by the MDMi cells. We propose that increased LRRK2 gene expression by MDMi cells could be a functional outcome of rs76904798, a single-nucleotide polymorphism in the LRKK2 locus that is associated with Parkinson's disease.
Copyright © 2017 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
References
-
- Bradshaw EM, Chibnik LB, Keenan BT, Ottoboni L, Raj T, Tang A, Rosenkrantz LL, Imboywa S, Lee M, Von Korff A, Morris MC, Evans DA, Johnson K, Sperling RA, Schneider JA, Bennett DA, De Jager PL. CD33 Alzheimer’s disease locus: altered monocyte function and amyloid biology. Nature neuroscience. 2013;16:848–850. - PMC - PubMed
-
- Guerreiro RJ, Lohmann E, Bras JM, Gibbs JR, Rohrer JD, Gurunlian N, Dursun B, Bilgic B, Hanagasi H, Gurvit H, Emre M, Singleton A, Hardy J. Using exome sequencing to reveal mutations in TREM2 presenting as a frontotemporal dementia-like syndrome without bone involvement. JAMA neurology. 2013;70:78–84. - PMC - PubMed
-
- Guerreiro R, Wojtas A, Bras J, Carrasquillo M, Rogaeva E, Majounie E, Cruchaga C, Sassi C, Kauwe JS, Younkin S, Hazrati L, Collinge J, Pocock J, Lashley T, Williams J, Lambert JC, Amouyel P, Goate A, Rademakers R, Morgan K, Powell J, St George-Hyslop P, Singleton A, Hardy J. TREM2 variants in Alzheimer’s disease. The New England journal of medicine. 2013;368:117–127. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
