

Identification of Residues Controlling Restriction versus Enhancing Activities of IFITM Proteins on Entry of Human Coronaviruses
- PMID: 29263263
- PMCID: PMC5827390
- DOI: 10.1128/JVI.01535-17
Identification of Residues Controlling Restriction versus Enhancing Activities of IFITM Proteins on Entry of Human Coronaviruses
Abstract
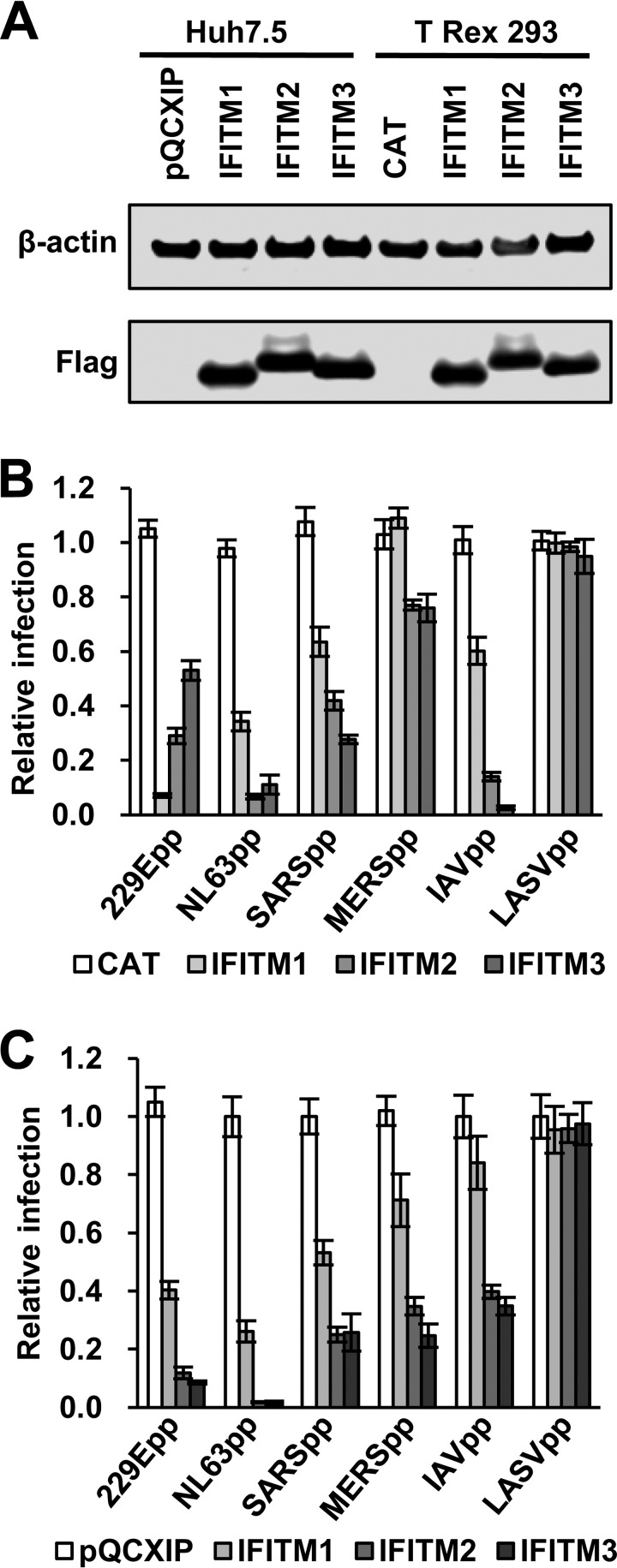
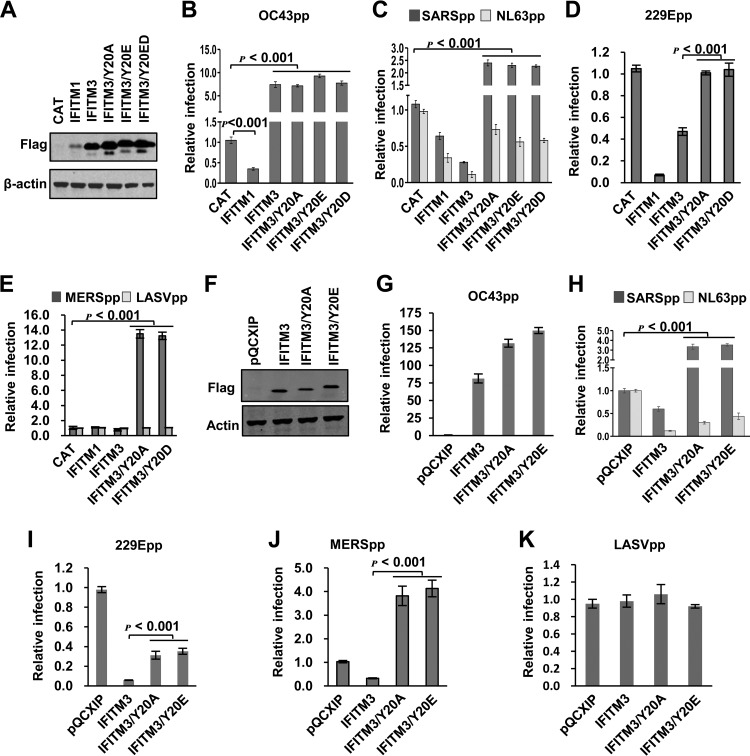
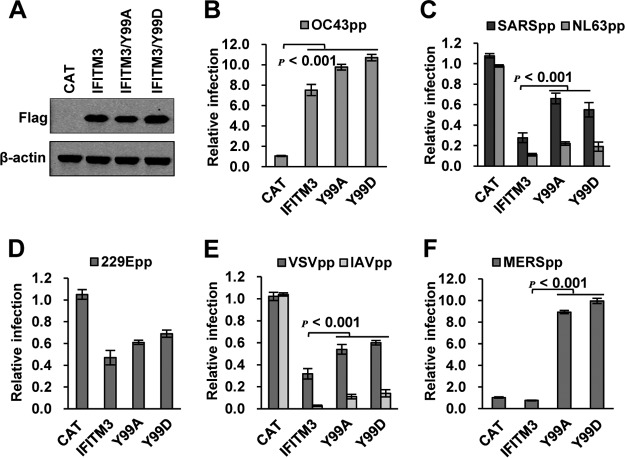
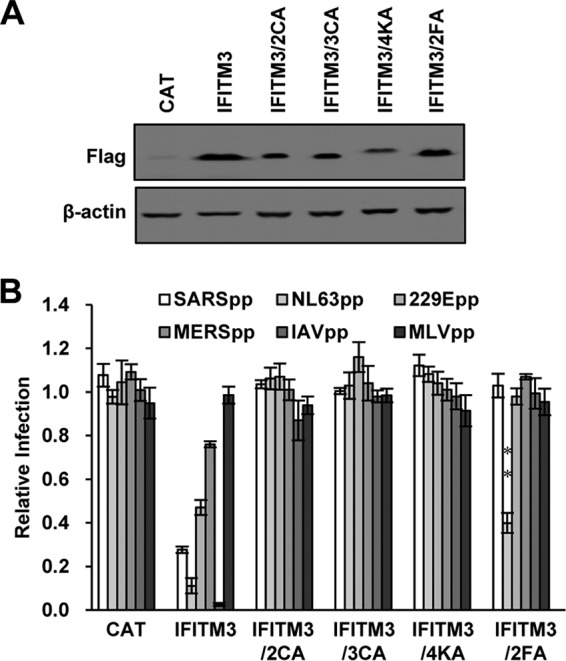
Interferon-induced transmembrane proteins (IFITMs) are restriction factors that inhibit the infectious entry of many enveloped RNA viruses. However, we demonstrated previously that human IFITM2 and IFITM3 are essential host factors facilitating the entry of human coronavirus (HCoV) OC43. In a continuing effort to decipher the molecular mechanism underlying IFITM differential modulation of HCoV entry, we investigated the roles of structural motifs important for IFITM protein posttranslational modifications, intracellular trafficking, and oligomerization in modulating the entry of five HCoVs. We found that three distinct mutations in IFITM1 or IFITM3 converted the host restriction factors to enhance entry driven by the spike proteins of severe acute respiratory syndrome coronavirus (SARS-CoV) and/or Middle East respiratory syndrome coronavirus (MERS-CoV). First, replacement of IFITM3 tyrosine 20 with either alanine or aspartic acid to mimic unphosphorylated or phosphorylated IFITM3 reduced its activity to inhibit the entry of HCoV-NL63 and -229E but enhanced the entry of SARS-CoV and MERS-CoV. Second, replacement of IFITM3 tyrosine 99 with either alanine or aspartic acid reduced its activity to inhibit the entry of HCoV-NL63 and SARS-CoV but promoted the entry of MERS-CoV. Third, deletion of the carboxyl-terminal 12 amino acid residues from IFITM1 enhanced the entry of MERS-CoV and HCoV-OC43. These findings suggest that these residues and structural motifs of IFITM proteins are key determinants for modulating the entry of HCoVs, most likely through interaction with viral and/or host cellular components at the site of viral entry to modulate the fusion of viral envelope and cellular membranes.IMPORTANCE The differential effects of IFITM proteins on the entry of HCoVs that utilize divergent entry pathways and membrane fusion mechanisms even when using the same receptor make the HCoVs a valuable system for comparative investigation of the molecular mechanisms underlying IFITM restriction or promotion of virus entry into host cells. Identification of three distinct mutations that converted IFITM1 or IFITM3 from inhibitors to enhancers of MERS-CoV or SARS-CoV spike protein-mediated entry revealed key structural motifs or residues determining the biological activities of IFITM proteins. These findings have thus paved the way for further identification of viral and host factors that interact with those structural motifs of IFITM proteins to differentially modulate the infectious entry of HCoVs.
Keywords: IFITM; coronavirus; viral entry.
Copyright © 2018 American Society for Microbiology.
Figures
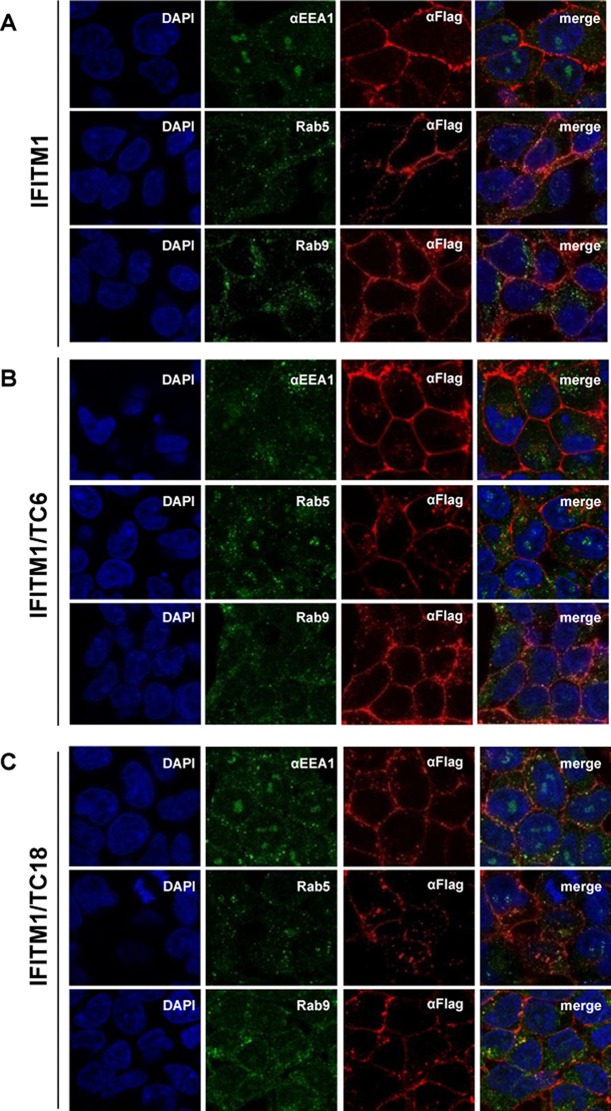
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
