

A Novel Ultrasensitive In Situ Hybridization Approach to Detect Short Sequences and Splice Variants with Cellular Resolution
- PMID: 29264769
- PMCID: PMC5994223
- DOI: 10.1007/s12035-017-0834-6
A Novel Ultrasensitive In Situ Hybridization Approach to Detect Short Sequences and Splice Variants with Cellular Resolution
Abstract
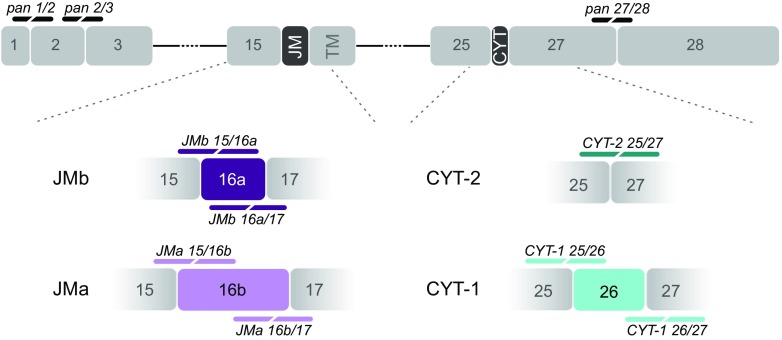
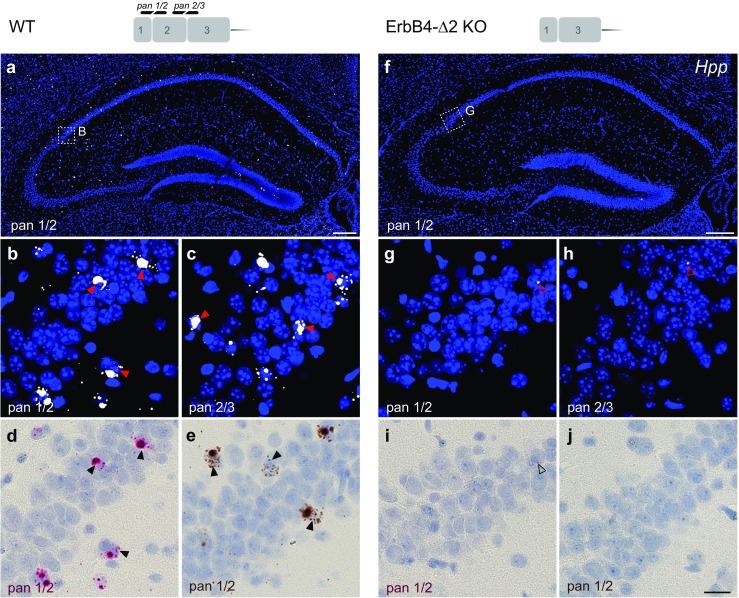
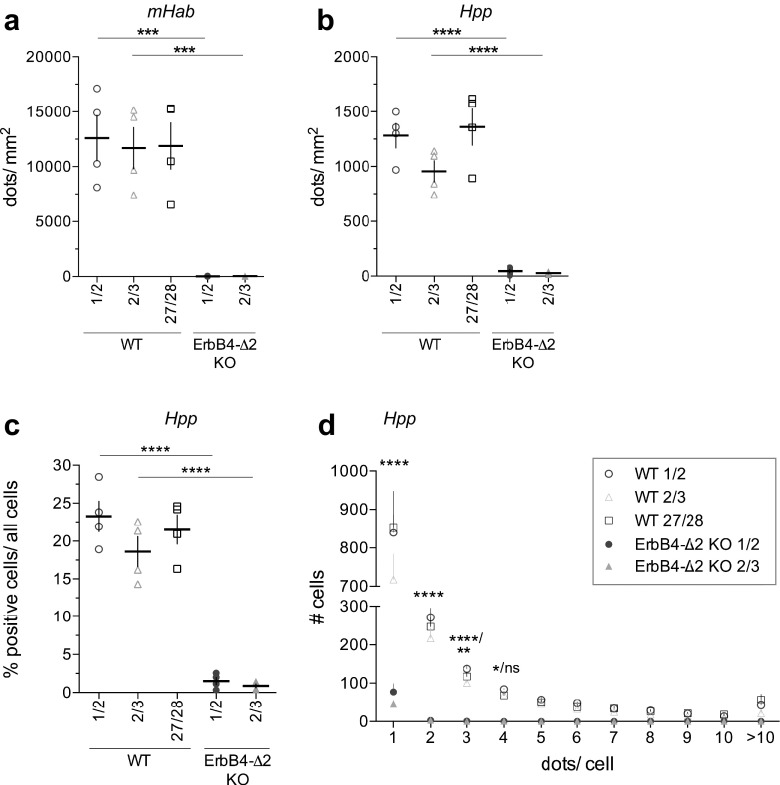
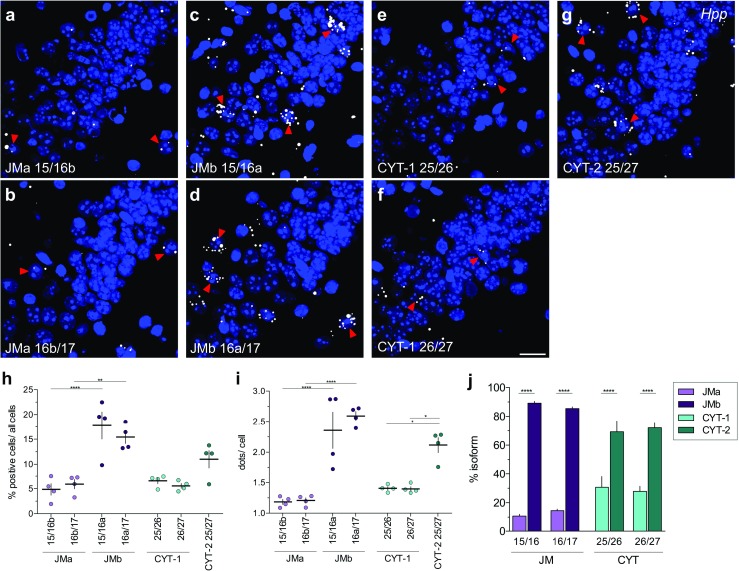
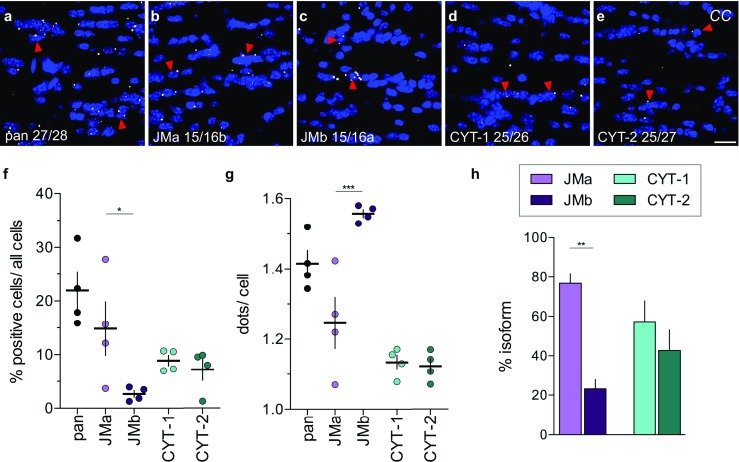
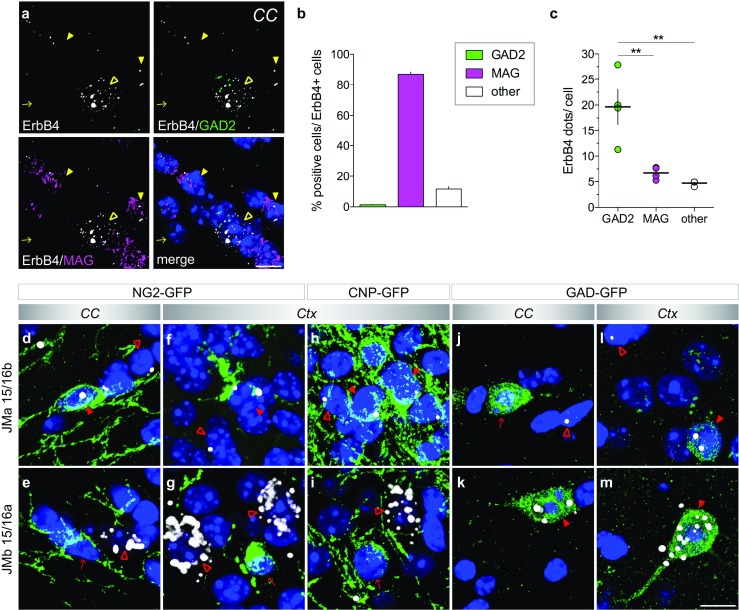
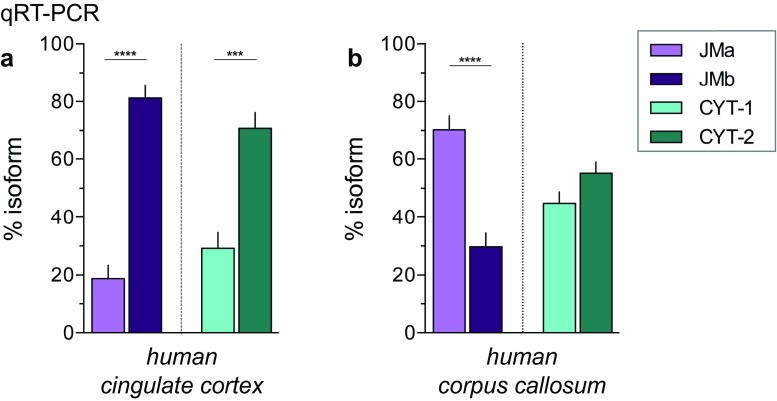
Investigating the expression of RNAs that differ by short or single nucleotide sequences at a single-cell level in tissue has been limited by the sensitivity and specificity of in situ hybridization (ISH) techniques. Detection of short isoform-specific sequences requires RNA isolation for PCR analysis-an approach that loses the regional and cell-type-specific distribution of isoforms. Having the capability to distinguish the differential expression of RNA variants in tissue is critical because alterations in mRNA splicing and editing, as well as coding single nucleotide polymorphisms, have been associated with numerous cancers, neurological and psychiatric disorders. Here we introduce a novel highly sensitive single-probe colorimetric/fluorescent ISH approach that targets short exon/exon RNA splice junctions using single-pair oligonucleotide probes (~ 50 bp). We use this approach to investigate, with single-cell resolution, the expression of four transcripts encoding the neuregulin (NRG) receptor ErbB4 that differ by alternative splicing of exons encoding two juxtamembrane (JMa/JMb) and two cytoplasmic (CYT-1/CYT-2) domains that alter receptor stability and signaling modes, respectively. By comparing ErbB4 hybridization on sections from wild-type and ErbB4 knockout mice (missing exon 2), we initially demonstrate that single-pair probes provide the sensitivity and specificity to visualize and quantify the differential expression of ErbB4 isoforms. Using cell-type-specific GFP reporter mice, we go on to demonstrate that expression of ErbB4 isoforms differs between neurons and oligodendrocytes, and that this differential expression of ErbB4 isoforms is evolutionarily conserved to humans. This single-pair probe ISH approach, known as BaseScope, could serve as an invaluable diagnostic tool to detect alternative spliced isoforms, and potentially single base polymorphisms, associated with disease.
Keywords: Alternative splicing; BaseScope; ErbB4; Neuregulin; Oligodendrocytes; RNA expression; Schizophrenia; Transcriptome.
Conflict of interest statement
L. E. and A.B. declare no competing financial interests. MX.H., A.L. and E.P. are employed by Advanced Cell Diagnostics.
Figures
References
-
- Kang HJ, Kawasawa YI, Cheng F, Zhu Y, Xu X, Li M, Sousa AM, Pletikos M, Meyer KA, Sedmak G, Guennel T, Shin Y, Johnson MB, Krsnik Z, Mayer S, Fertuzinhos S, Umlauf S, Lisgo SN, Vortmeyer A, Weinberger DR, Mane S, Hyde TM, Huttner A, Reimers M, Kleinman JE, Sestan N. Spatio-temporal transcriptome of the human brain. Nature. 2011;478(7370):483–489. doi: 10.1038/nature10523. - DOI - PMC - PubMed
-
- Zhang Y, Chen K, Sloan SA, Bennett ML, Scholze AR, O'Keeffe S, Phatnani HP, Guarnieri P, Caneda C, Ruderisch N, Deng S, Liddelow SA, Zhang C, Daneman R, Maniatis T, Barres BA, Wu JQ. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J Neurosci. 2014;34(36):11929–11947. doi: 10.1523/JNEUROSCI.1860-14.2014. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
