

The role of phospholipid molecular species in determining the physical properties of yeast membranes
- PMID: 29265372
- PMCID: PMC5947837
- DOI: 10.1002/1873-3468.12944
The role of phospholipid molecular species in determining the physical properties of yeast membranes
Abstract
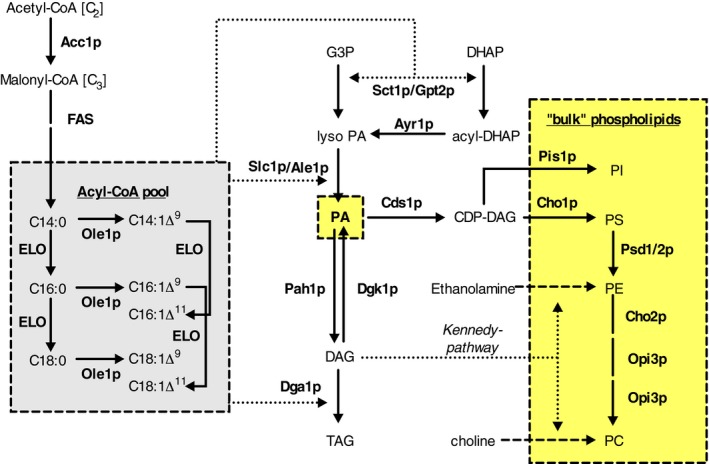
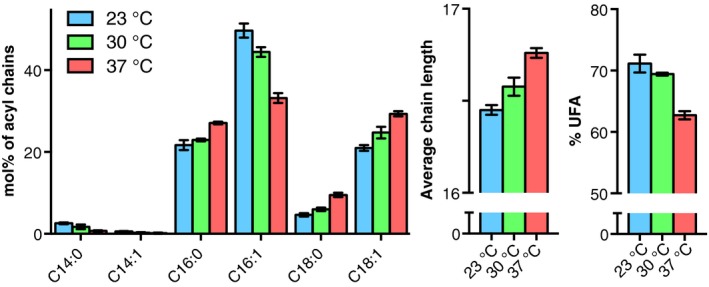
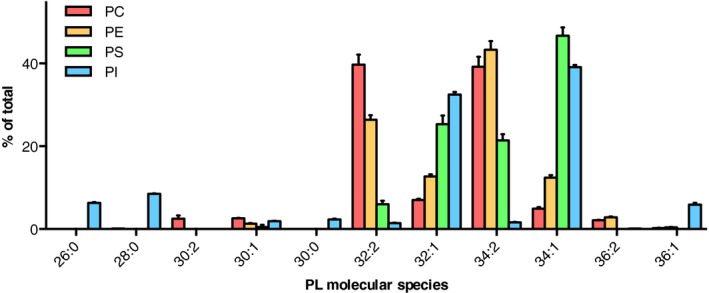
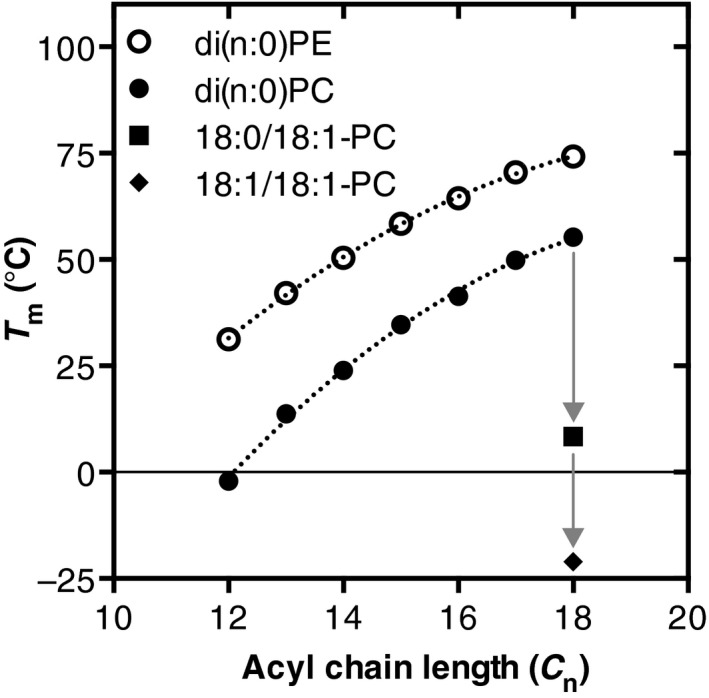
In most eukaryotes, including Saccharomyces cerevisiae, glycerophospholipids are the main membrane lipid constituents. Besides serving as general membrane 'building blocks', glycerophospholipids play an important role in determining the physical properties of the membrane, which are crucial for proper membrane function. To ensure optimal physical properties, membrane glycerophospholipid composition and synthesis are tightly regulated. This review will summarize our current knowledge of factors and processes determining the membrane glycerophospholipid composition of the reference eukaryote S. cerevisiae at the level of molecular species. Extrapolating from relevant model membrane data, we also discuss how modulation of the molecular species composition can regulate membrane physical properties.
Keywords: membrane fluidity; phospholipid molecular species; phospholipid properties.
© 2017 The Authors. FEBS Letters published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Figures
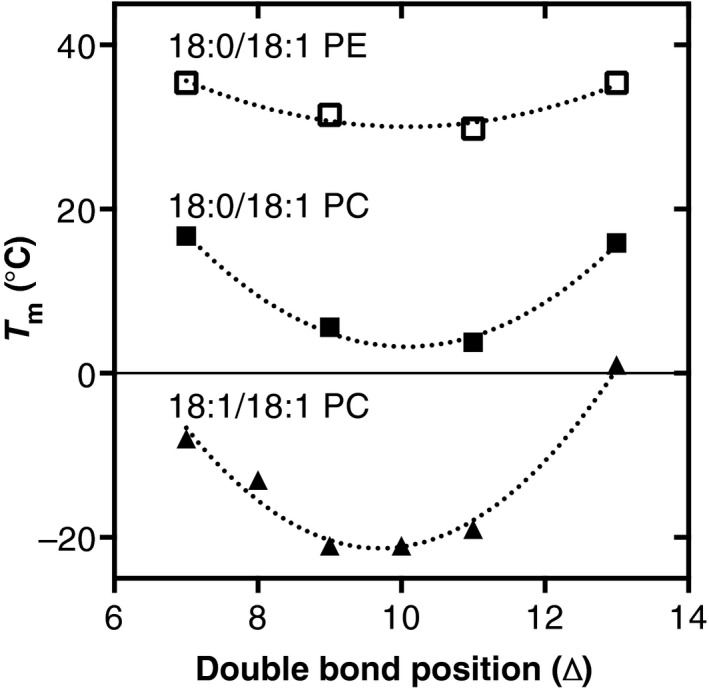
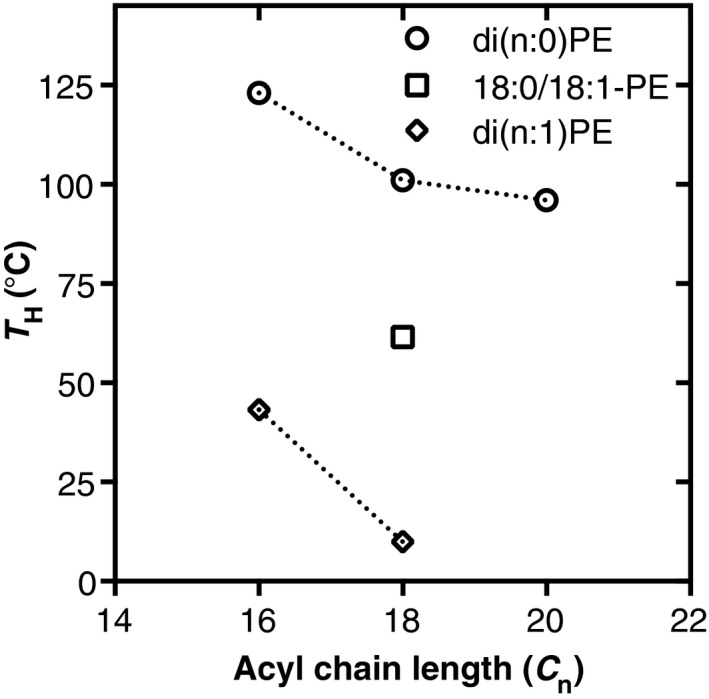
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
