

Outer membrane protein folding from an energy landscape perspective
- PMID: 29268734
- PMCID: PMC5740924
- DOI: 10.1186/s12915-017-0464-5
Outer membrane protein folding from an energy landscape perspective
Abstract
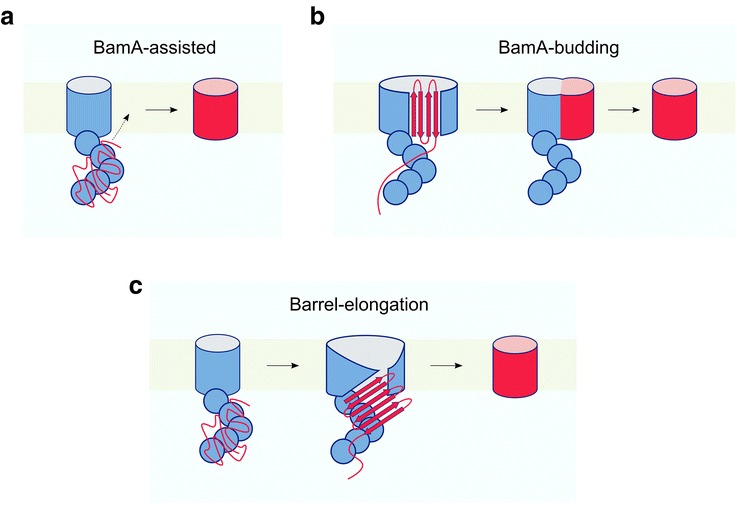
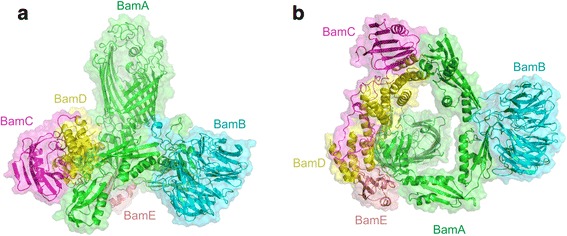
The cell envelope is essential for the survival of Gram-negative bacteria. This specialised membrane is densely packed with outer membrane proteins (OMPs), which perform a variety of functions. How OMPs fold into this crowded environment remains an open question. Here, we review current knowledge about OMP folding mechanisms in vitro and discuss how the need to fold to a stable native state has shaped their folding energy landscapes. We also highlight the role of chaperones and the β-barrel assembly machinery (BAM) in assisting OMP folding in vivo and discuss proposed mechanisms by which this fascinating machinery may catalyse OMP folding.
Conflict of interest statement
Competing interests
The authors have no financial and non-financial competing interests.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
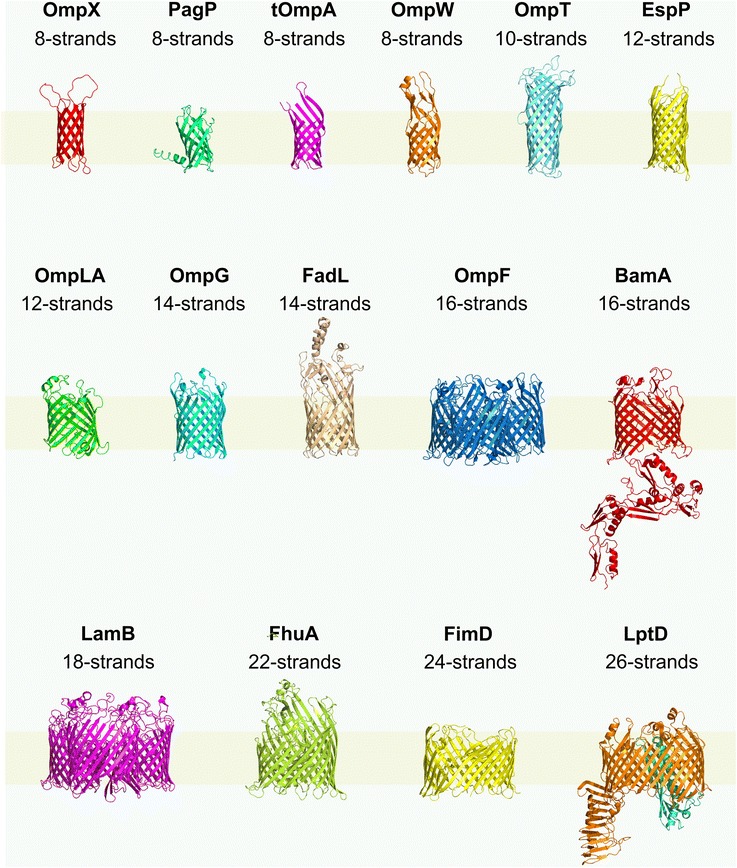
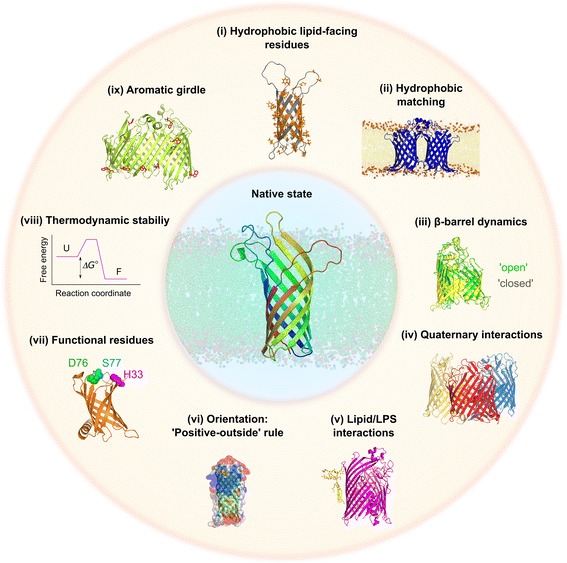
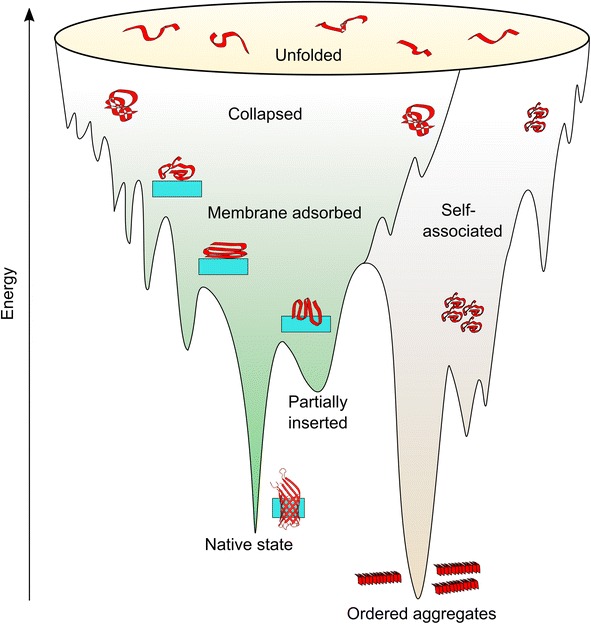
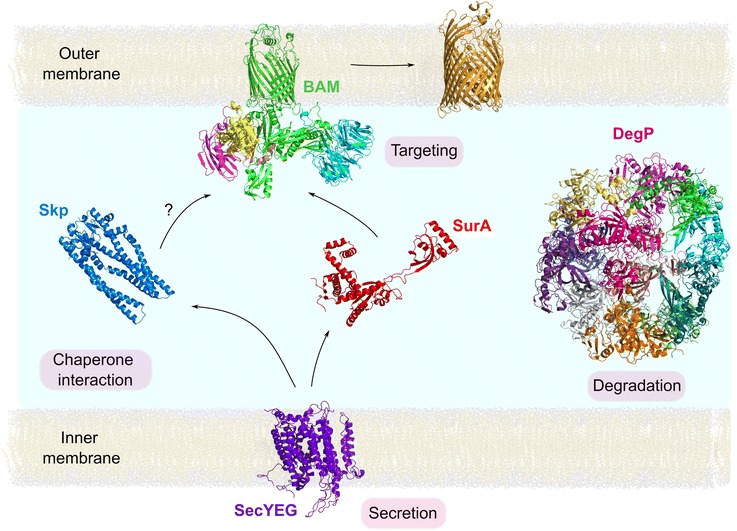
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
