

Plant Lipid Droplets and Their Associated Proteins: Potential for Rapid Advances
- PMID: 29269574
- PMCID: PMC5841732
- DOI: 10.1104/pp.17.01677
Plant Lipid Droplets and Their Associated Proteins: Potential for Rapid Advances
Abstract
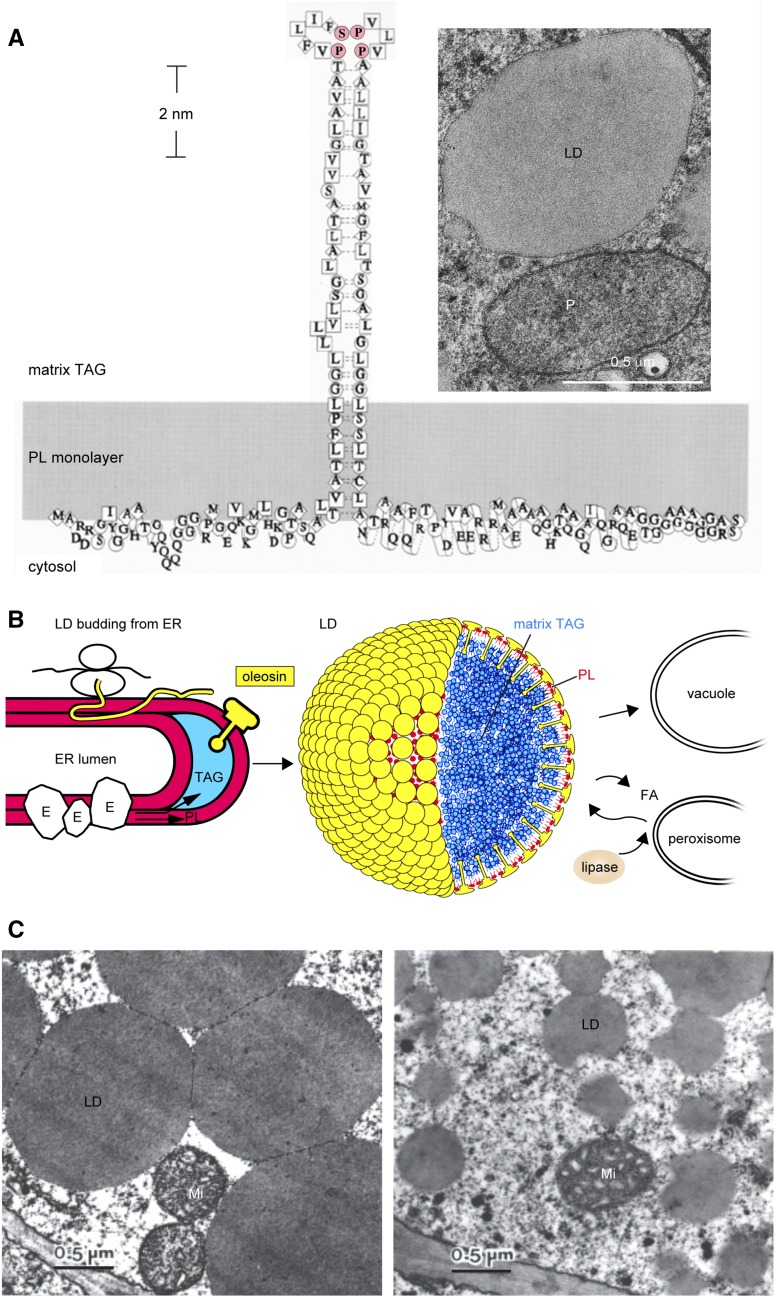
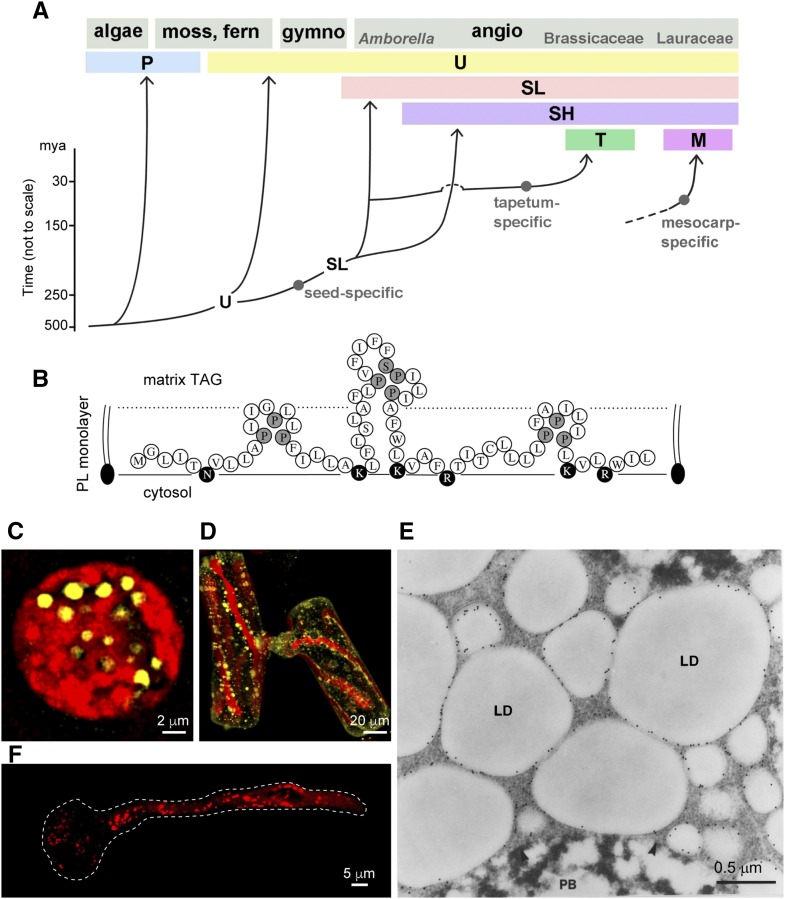
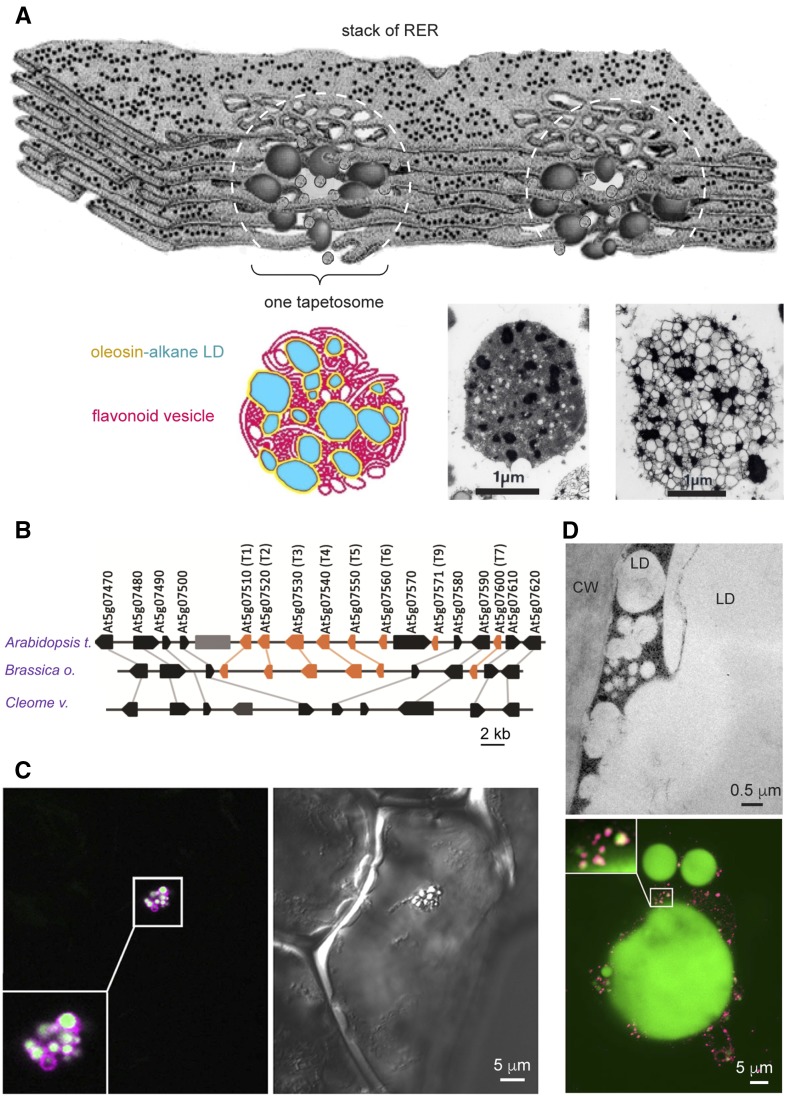
Cytoplasmic lipid droplets (LDs) of neutral lipids (triacylglycerols [TAGs], sterylesters, etc.) are reserves of high-energy metabolites and other constituents for future needs. They are present in diverse cells of eukaryotes and prokaryotes. An LD has a core of neutral lipids enclosed with a monolayer of phospholipids and proteins, which play structural and/or metabolic roles. During the past 3 decades, studies of LDs in diverse organisms have blossomed after they were found to be involved in prevalent human diseases and industrial uses. LDs in plant seeds were studied before those in mammals and microbes, and the latter studies have since moved forward. Plant LDs carry a hallmark protein called oleosin, which has a long hydrophobic hairpin penetrating the TAG core and stabilizing the LD. The oleosin gene first appeared in green algae and has evolved in enhancing promoter strength, tandem repeats, and/or expression specificity, leading to the appearance of new LD organelles, such as tapetosomes in Brassicaceae. The synthesis of LDs occurs with TAG-synthesizing enzymes on the endoplasmic reticulum (ER), and nascent TAGs are sequestered in the acyl moiety region between the bilayers of phospholipids, which results in ER-LD swelling. Oleosin is synthesized on the cytosol side of the ER and extracts the LD from the ER-LD to cytosol. This extraction of LD to the cytosol is controlled solely by the innate properties of oleosin, and modified oleosin can redirect the LD to the ER lumen and then vacuoles. The breakdown of LDs requires lipase associating with core retromer and binding to peroxisomes, which then send the enzyme to LDs via tubular extensions. Two groups of LD-associated proteins, caleosin/dioxygenase/steroleosin and LD/oil body-associated proteins, participate in cellular stress defenses via enzymic activities and binding, respectively. The surface of LDs in all plant cells may be an inert refuge for these and other proteins, which exert functions on diverse cell components. Oleosin-LDs have been explored for commercial applications; successes in their uses will rely on overcoming conceptual and technical difficulties.
© 2018 American Society of Plant Biologists. All Rights Reserved.
Figures


References
-
- Abell BM, Hahn M, Holbrook LA, Moloney MM (2004) Membrane topology and sequence requirements for oil body targeting of oleosin. Plant J 37: 461–470 - PubMed
-
- Abell BM, High S, Moloney MM (2002) Membrane protein topology of oleosin is constrained by its long hydrophobic domain. J Biol Chem 277: 8602–8610 - PubMed
-
- Alameldin H, Izadi-Darbandi A, Smith SA, Balan V, Jonese AD, Orhun GE, Sticklen M (2017) Metabolic engineering to increase the corn seed storage lipid quantity and change its compositional quality. Crop Sci 57: 1854–1864
-
- Alexander LG, Sessions RB, Clarke AR, Tatham AS, Shewry PR, Napier JA (2002) Characterization and modelling of the hydrophobic domain of a sunflower oleosin. Planta 214: 546–551 - PubMed
-
- Barbosa AD, Siniossoglou S (2017) Function of lipid droplet-organelle interactions in lipid homeostasis. Biochim Biophys Acta 1864: 1459–1468 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
