

Review
doi: 10.1038/s41556-017-0012-0.
Epub 2017 Dec 21.
Mechanoreciprocity in cell migration
Affiliations
- PMID: 29269951
- PMCID: PMC5943039
- DOI: 10.1038/s41556-017-0012-0
Item in Clipboard
Review
Mechanoreciprocity in cell migration
Nat Cell Biol.
2018 Jan.
Abstract
Cell migration is an adaptive process that depends on and responds to physical and molecular triggers. Moving cells sense and respond to tissue mechanics and induce transient or permanent tissue modifications, including extracellular matrix stiffening, compression and deformation, protein unfolding, proteolytic remodelling and jamming transitions. Here we discuss how the bi-directional relationship of cell-tissue interactions (mechanoreciprocity) allows cells to change position and contributes to single-cell and collective movement, structural and molecular tissue organization, and cell fate decisions.
Figures
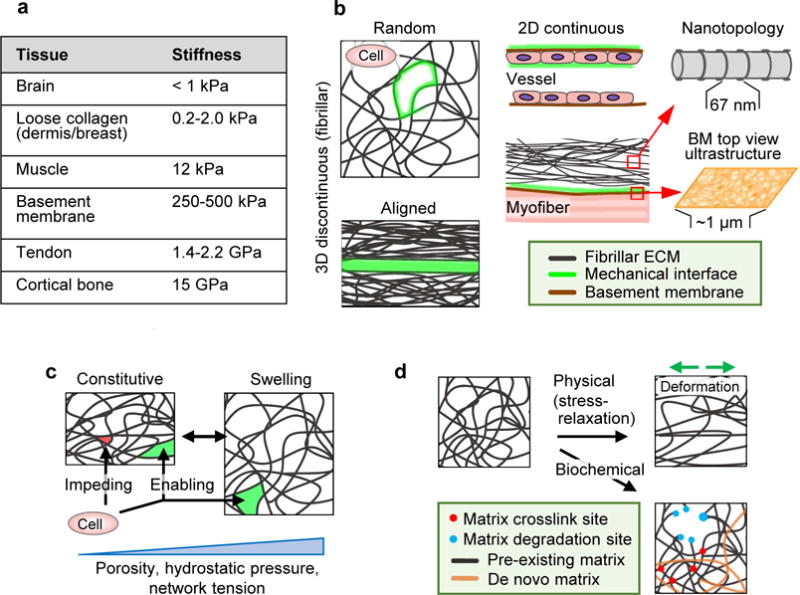
Tissue properties determining cell migration, including topology of the ECM and organization of interfaces between tissue structures. A Elastic modulus range present in macromolecular tissue structures. GAG-rich stroma surrounding cell networks, such as brain tissue, is soft (below 1.0 kPa); loose fibrillar type I and III collagen-based porous protein networks such as dermis and breast scale typically between 0.2 and 2 kPa,. Thicker, more crosslinked collagen bundles in muscle are substantially stiffer (12 kPa), which reaches the low GPa range in tendon. Basement membranes consisting of type IV collagen and laminins have a stiffness in the higher kPa range, with at least double the stiffness on the epithelial side compared to the stromal side. The stiffness of calcified tissue, including cortical bone, can go up to 15 GPa. B Principal ECM geometries defining mechanical cell migration interfaces (green) including 1D, 2D and 3D organization and nanotopology. C Baseline porosity range, relative to cell size and deformability, and hydrostatic pressure induced reversible swelling and mechanical alterations in fibrillar ECM. D Irreversible changes induced by physical or biochemical factors. Direction of physical deformation denoted by green arrows.
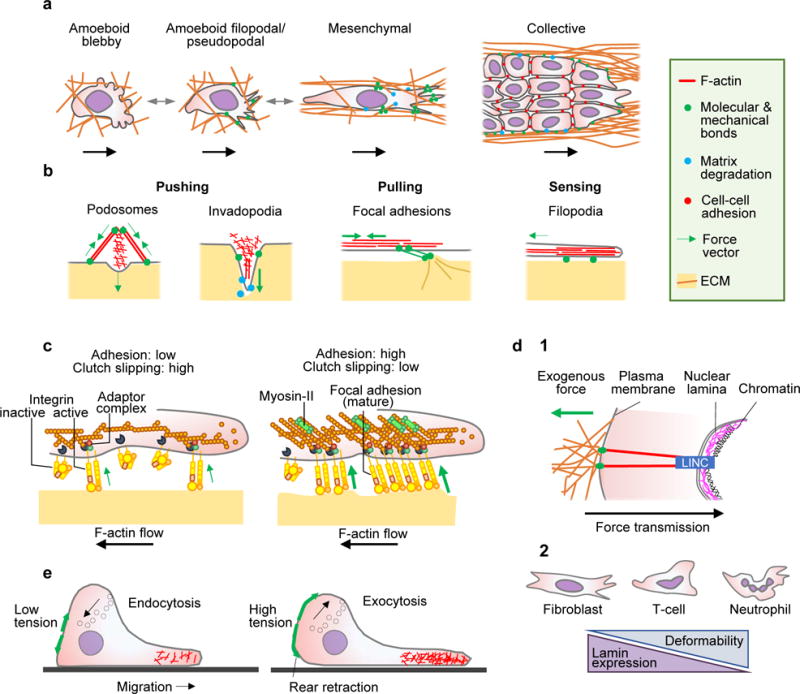
A Cell migration modes in 3D environments, including single-cell and collective migration. B Actin-rich cell surface structures mechanically interacting with tissue, including: podosomes probing the substrate vertically; invadopodia protruding and locally degrading substrate in vertical direction by releasing proteases; focal adhesions generating adhesion and pulling force; and filopodia extending along ECM structures and generating traction force in parallel. C Differential adhesion regulation. Tension generated by mechanical extracellular interactions of adhesion receptors depend upon intracellular adaptor proteins linking to the actin cytoskeleton, which flows in rearward direction (clutch). The strength and duration of adhesion receptor bonds to substrate increase with force and are regulated by lateral clustering of integrin adhesion receptors, which increases the number of bonds. Diffusely distributed integrins exert weak adhesion and traction force (left). With integrin clustering, adhesions increase the number of engaged bonds to actin filaments and recruit myosin-II, providing stronger adhesion and traction force towards the substrate (right). D Nuclear mechanics. (1) Mechanical linkage between ECM, cytoskeleton, nuclear lamina and chromatin through the LINC complex (linker of nucleoskeleton and cytoskeleton) consisting of nesprins 1-4 and Sun1/2 proteins, which regulate nuclear positioning and deformation in response to cell responses to extracellular cues. (2) Shapes and correlation between nuclear deformability and lamin expression. E Membrane tension and cell migration. Left panel, low membrane tension in partly polarized cell facilitating actin polymerization. Right panel, fully polarized cell with high membrane tension limits actin filament protrusion at the leading edge, but supports rear contraction and is counterbalanced by transport of intracellular vesicles to the plasma membrane. Green arrows, force vectors.

A (l) Spiral concept of mechanical cell-tissue interactions, describing coevolution of cell and tissue mechanics. Cells impose ‘mediators’ (pulling, pushing, ECM deposition, ECM degradation) and thereby alter tissue modules. Through iterative reinforcement (indicated by the spiral) or negative feedback, both cell and tissue modules undergo coevolution towards altered morphology and function. (2) Development of cell and tissue coevolution, including acquisition of a stable equilibrium (left), oscillatory coevolution with both positive and negative feedback loops (middle), or unstable coevolution, typically caused by negative feedback or disruption of co-engagement (right). The dashed lines indicate the temporal coevolution for perturbed initial conditions; in case of stable or oscillatory coevolution, the system converges to well-defined asymptotic behaviour, in the case of unstable coevolution the sensitive dependence on initial conditions typical of chaotic systems is seen. B Mechanoreciprocity in cell migration. (1) Strain stiffening of an ECM network by tension at the leading edge, creating a multi-parameter mechanochemical feed-forward loop. (2) Tissue compression by migrating cells, condensing ECM. Intracellular hydrostatic pressure is jointly maintained by stress-sensitive ion and water channels and actomyosin contractility. (3) Structural ECM remodelling. Mesenchymal migration leads to traction and fibre realignment by the leading edge, followed by pericellular proteolysis of collagen fibrils constraining the cell body, which after fibril realignment leaves behind a remodelled tissue track. C Molecular mechanoreciprocity. (1) Unfolding of mechanosensitive proteins by force. Actomyosin-contraction and tension exposes bioactive domains in adaptor proteins (e.g., talin, vinculin, p130CAS), which allows further ligands to bind and alter function in a strain-dependent manner (e.g. increased lifetime of catch-bonds). (2) Strain-dependent activation of TGF-β1. Cell adhesion and tension to ECM-tethered LAP induces a conformational change and releases TGF-β to diffuse and bind to its receptor. LTBP-1, latent TGF-β-binding protein 1. D Mechanoreciprociticy in disease. (1) Concordant immigration of fibroblasts and endothelial cells into the wound bed, which realign and degrade provisional ECM and synthesize collagen and basement membrane proteins and undergo a transition of engaged integrin systems. As outcomes, tissue alignment, density and stiffness are reciprocally linked to fibroblast function. (2) Mechanoreciprocity in cancer invasion. Dual function of ECM deposition and stiffening by myofibroblasts in sub regions, leading to encapsulation or invasion along collagen interfaces.
References
-
- Charras G, Sahai E. Physical influences of the extracellular environment on cell migration. Nat Rev Mol Cell Biol. 2014;15:813–824. - PubMed
-
- Te Boekhorst V, Preziosi L, Friedl P. Plasticity of Cell Migration In Vivo and In Silico. Annu Rev Cell Dev Biol. 2016;3228361:1–28. - PubMed
-
- Bornstein P, McPherson J, Sage H. In: Pathobiology of the endothelial cell. Nossel H, Vogel H, editors. New York: Academic Press; 1982. pp. 215–228.
-
- Bissell MJ, Hall HG, Parry G. How does the extracellular matrix direct gene expression? J Theor Biol. 1982;99:31–68. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
