

Parallel evolution of the POQR prolyl oligo peptidase gene conferring plant quantitative disease resistance
- PMID: 29272270
- PMCID: PMC5757927
- DOI: 10.1371/journal.pgen.1007143
Parallel evolution of the POQR prolyl oligo peptidase gene conferring plant quantitative disease resistance
Abstract
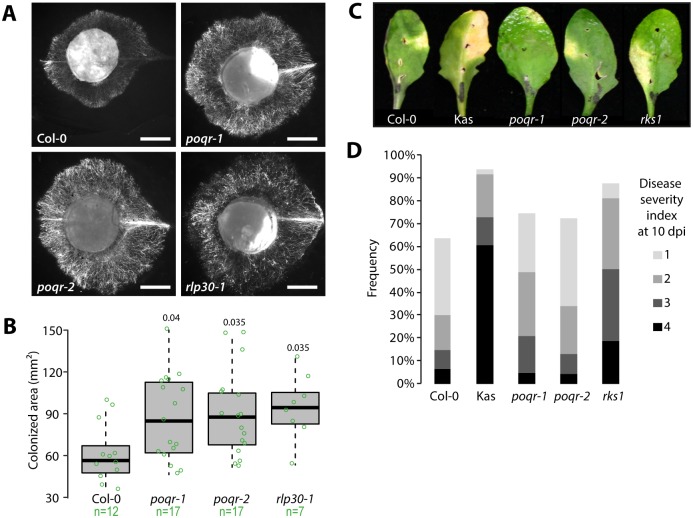
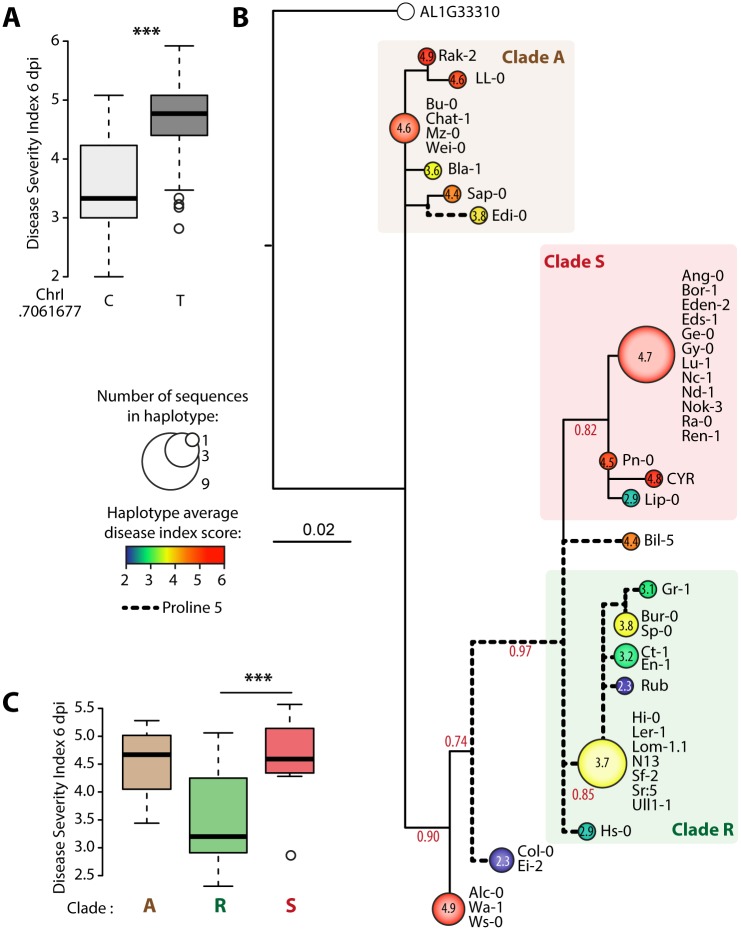
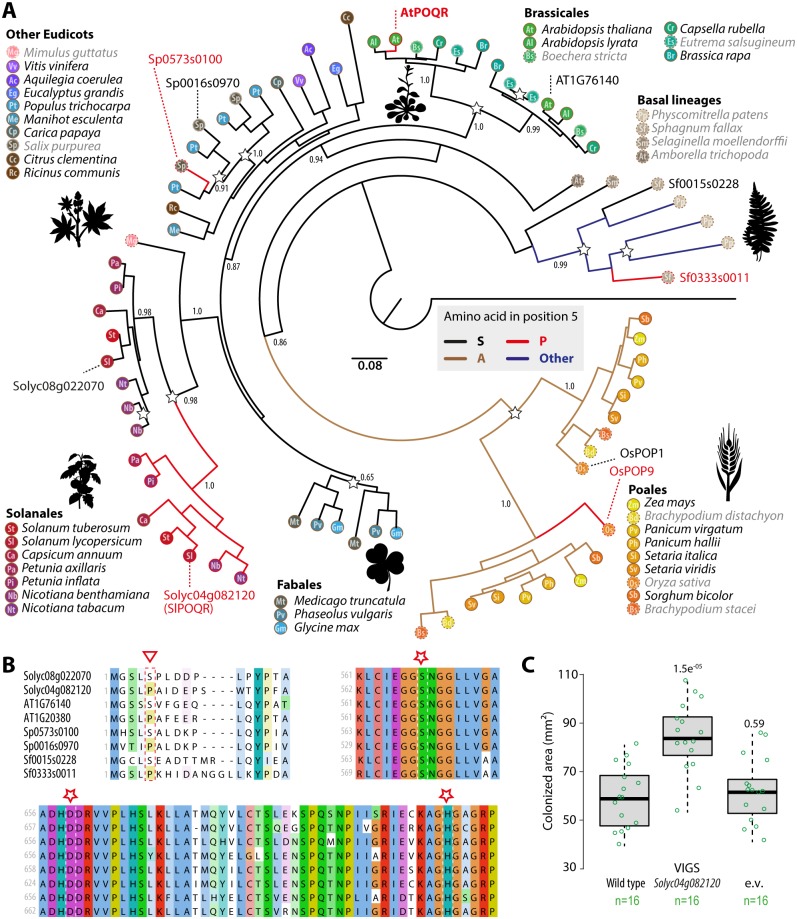
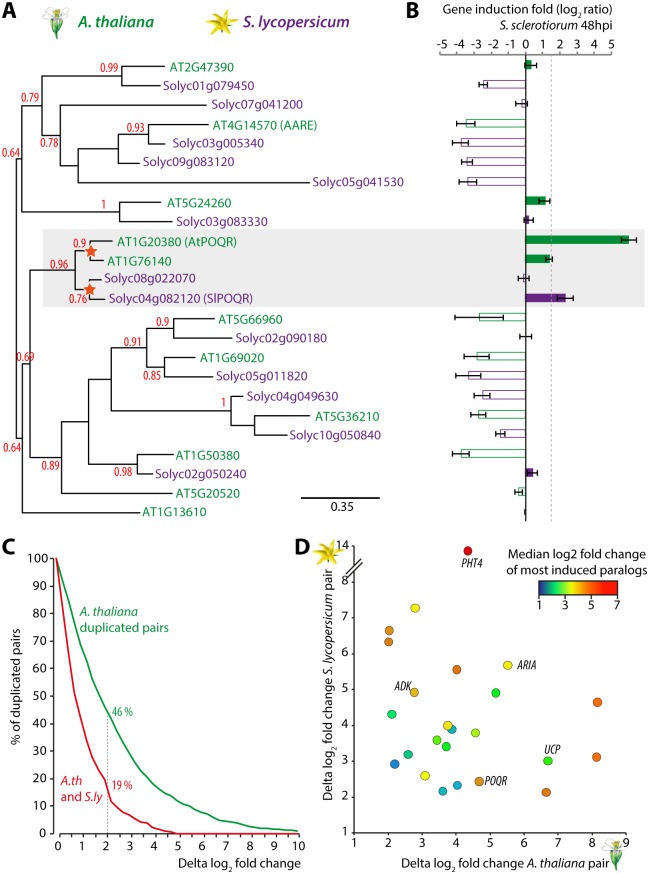
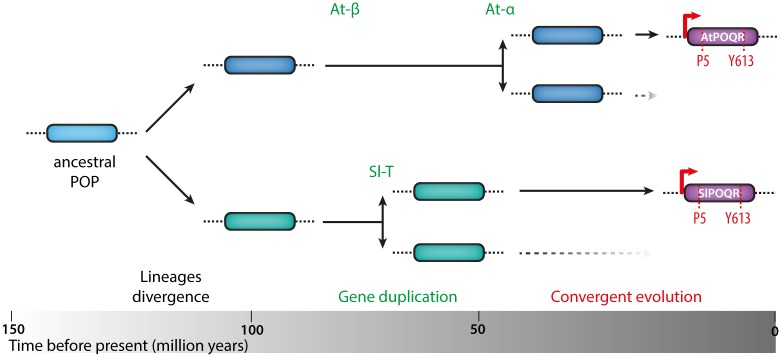
Plant pathogens with a broad host range are able to infect plant lineages that diverged over 100 million years ago. They exert similar and recurring constraints on the evolution of unrelated plant populations. Plants generally respond with quantitative disease resistance (QDR), a form of immunity relying on complex genetic determinants. In most cases, the molecular determinants of QDR and how they evolve is unknown. Here we identify in Arabidopsis thaliana a gene mediating QDR against Sclerotinia sclerotiorum, agent of the white mold disease, and provide evidence of its convergent evolution in multiple plant species. Using genome wide association mapping in A. thaliana, we associated the gene encoding the POQR prolyl-oligopeptidase with QDR against S. sclerotiorum. Loss of this gene compromised QDR against S. sclerotiorum but not against a bacterial pathogen. Natural diversity analysis associated POQR sequence with QDR. Remarkably, the same amino acid changes occurred after independent duplications of POQR in ancestors of multiple plant species, including A. thaliana and tomato. Genome-scale expression analyses revealed that parallel divergence in gene expression upon S. sclerotiorum infection is a frequent pattern in genes, such as POQR, that duplicated both in A. thaliana and tomato. Our study identifies a previously uncharacterized gene mediating QDR against S. sclerotiorum. It shows that some QDR determinants are conserved in distantly related plants and have emerged through the repeated use of similar genetic polymorphisms at different evolutionary time scales.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Dodds PN, Rathjen JP (2010) Plant immunity: towards an integrated view of plant–pathogen interactions. Nature Reviews Genetics 11: 539–548. doi: 10.1038/nrg2812 - DOI - PubMed
-
- Takken FL, Goverse A (2012) How to build a pathogen detector: structural basis of NB-LRR function. Current opinion in plant biology 15: 375–384. doi: 10.1016/j.pbi.2012.05.001 - DOI - PubMed
-
- Le Roux C, Huet G, Jauneau A, Camborde L, Trémousaygue D, et al. (2015) A receptor pair with an integrated decoy converts pathogen disabling of transcription factors to immunity. Cell 161: 1074–1088. doi: 10.1016/j.cell.2015.04.025 - DOI - PubMed
-
- Poland JA, Balint-Kurti PJ, Wisser RJ, Pratt RC, Nelson RJ (2009) Shades of gray: the world of quantitative disease resistance. Trends in plant science 14: 21–29. doi: 10.1016/j.tplants.2008.10.006 - DOI - PubMed
-
- Roux F, Voisin D, Badet T, Balagué C, Barlet X, et al. (2014) Resistance to phytopathogens e tutti quanti: placing plant quantitative disease resistance on the map. Molecular Plant Pathology 15: 427–432. doi: 10.1111/mpp.12138 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
