

Complex Relationships between Chromatin Accessibility, Sequence Divergence, and Gene Expression in Arabidopsis thaliana
- PMID: 29272536
- PMCID: PMC5889003
- DOI: 10.1093/molbev/msx326
Complex Relationships between Chromatin Accessibility, Sequence Divergence, and Gene Expression in Arabidopsis thaliana
Erratum in
-
Molecular Biology and Evolution, Volume 35, Issue 4.Mol Biol Evol. 2018 Jul 1;35(7):1821. doi: 10.1093/molbev/msy057. Mol Biol Evol. 2018. PMID: 29659984 Free PMC article. No abstract available.
Abstract
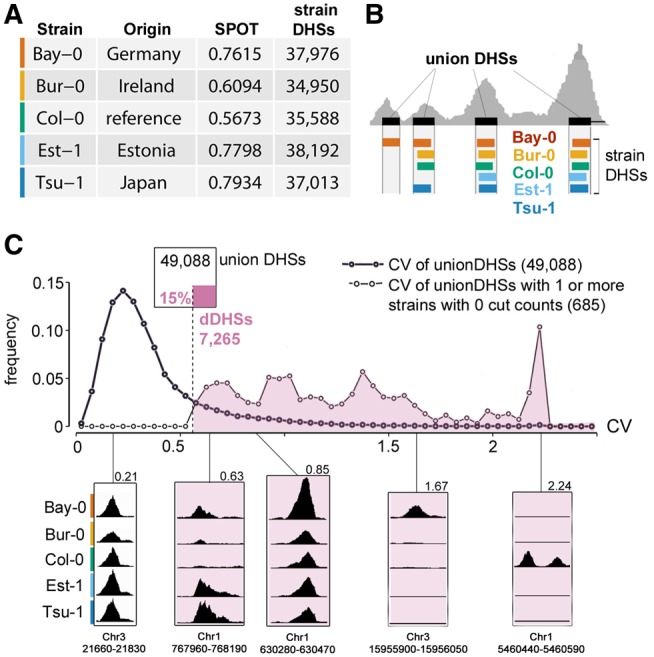
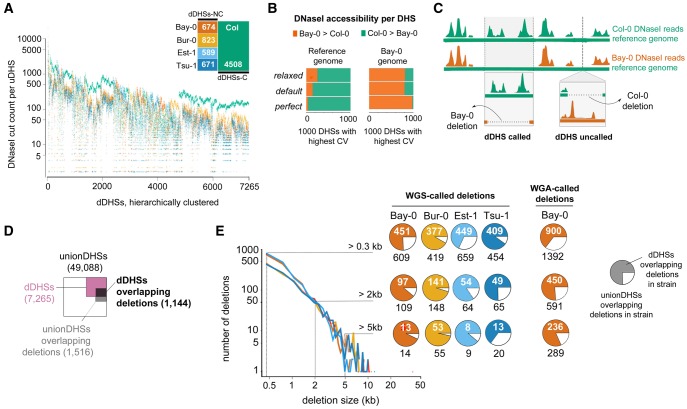
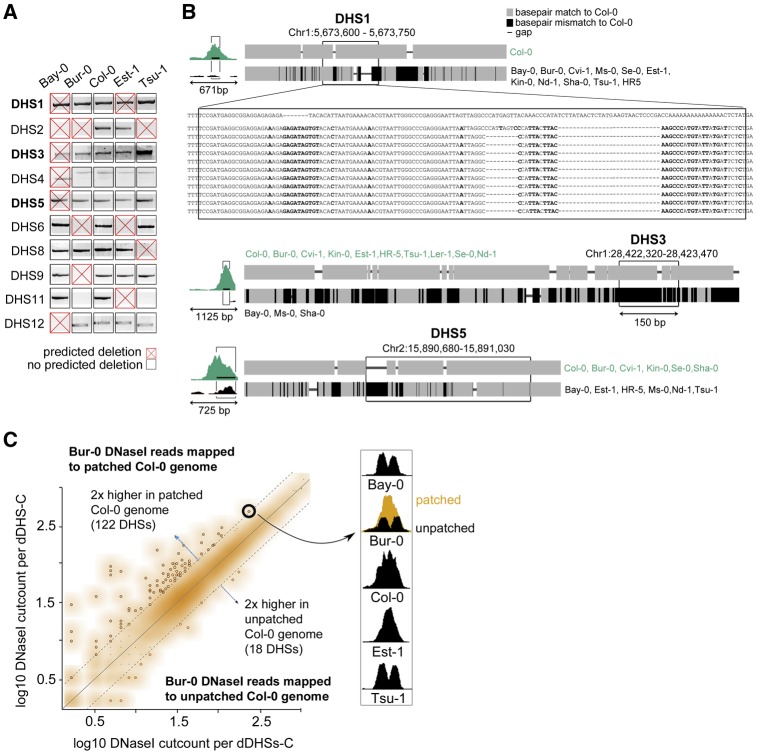
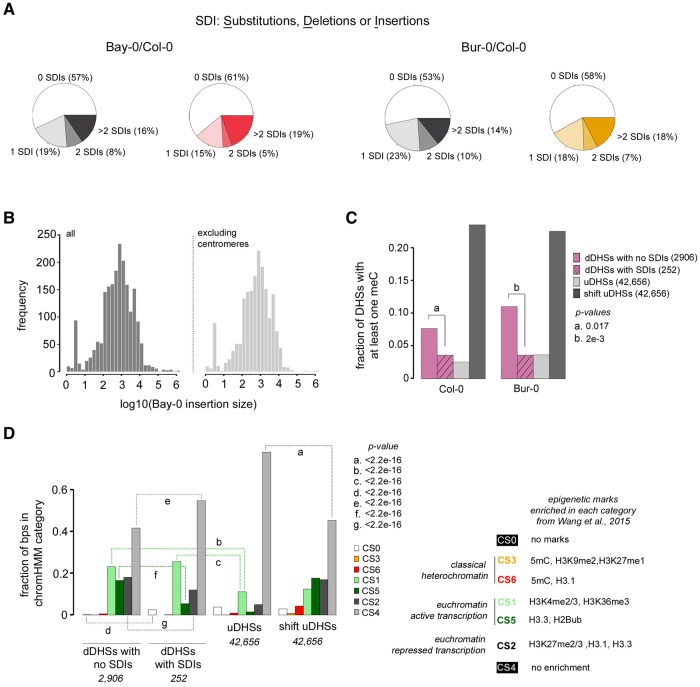
Variation in regulatory DNA is thought to drive phenotypic variation, evolution, and disease. Prior studies of regulatory DNA and transcription factors across animal species highlighted a fundamental conundrum: Transcription factor binding domains and cognate binding sites are conserved, while regulatory DNA sequences are not. It remains unclear how conserved transcription factors and dynamic regulatory sites produce conserved expression patterns across species. Here, we explore regulatory DNA variation and its functional consequences within Arabidopsis thaliana, using chromatin accessibility to delineate regulatory DNA genome-wide. Unlike in previous cross-species comparisons, the positional homology of regulatory DNA is maintained among A. thaliana ecotypes and less nucleotide divergence has occurred. Of the ∼50,000 regulatory sites in A. thaliana, we found that 15% varied in accessibility among ecotypes. Some of these accessibility differences were associated with extensive, previously unannotated sequence variation, encompassing many deletions and ancient hypervariable alleles. Unexpectedly, for the majority of such regulatory sites, nearby gene expression was unaffected. Nevertheless, regulatory sites with high levels of sequence variation and differential chromatin accessibility were the most likely to be associated with differential gene expression. Finally, and most surprising, we found that the vast majority of differentially accessible sites show no underlying sequence variation. We argue that these surprising results highlight the necessity to consider higher-order regulatory context in evaluating regulatory variation and predicting its phenotypic consequences.
Figures


References
-
- Arnold CD, Gerlach D, Stelzer C, Boryń ŁM, Rath M, Stark A.. 2013. Genome-wide quantitative enhancer activity maps identified by STARR-seq. Science 3396123:1074–1077. - PubMed
-
- Beadle George W. 1972. The Mystery of Maize. In Joyce Zibro, editor. Field Museum of Natural History Bulletin, p. 212–221.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
