

Transcriptional and functional profiling defines human small intestinal macrophage subsets
- PMID: 29273642
- PMCID: PMC5789404
- DOI: 10.1084/jem.20170057
Transcriptional and functional profiling defines human small intestinal macrophage subsets
Abstract
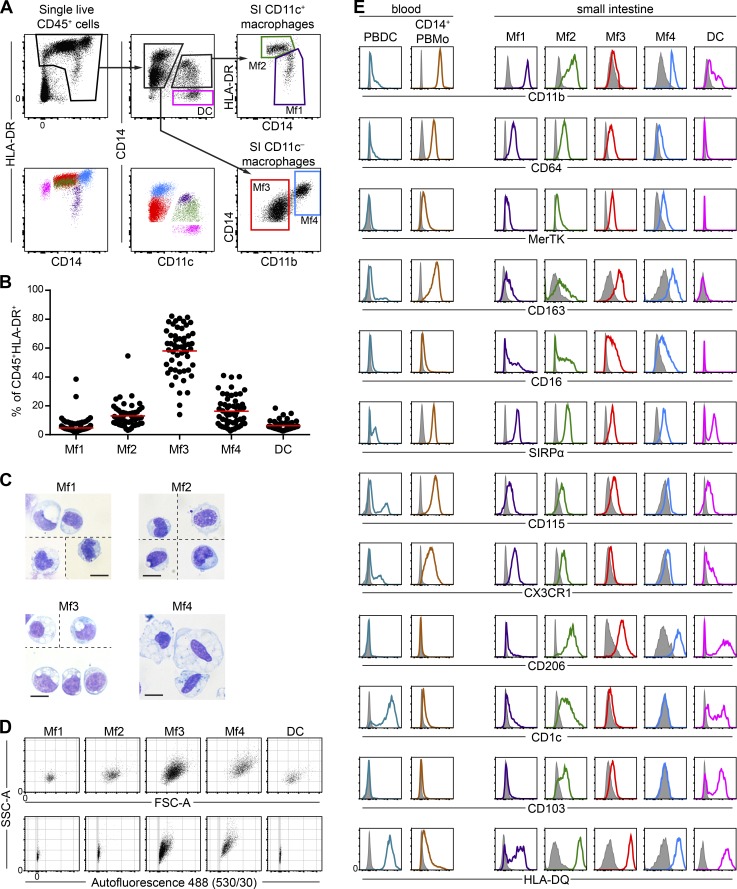
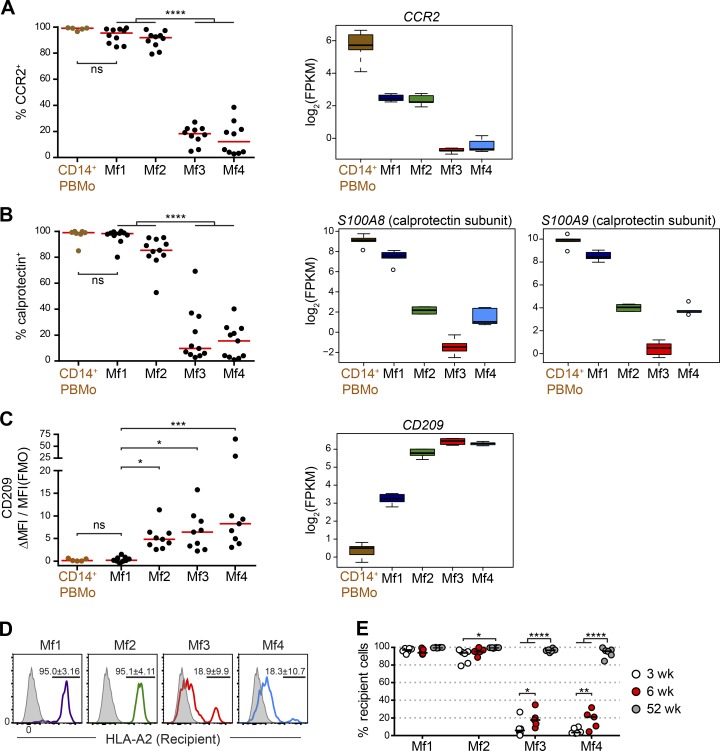
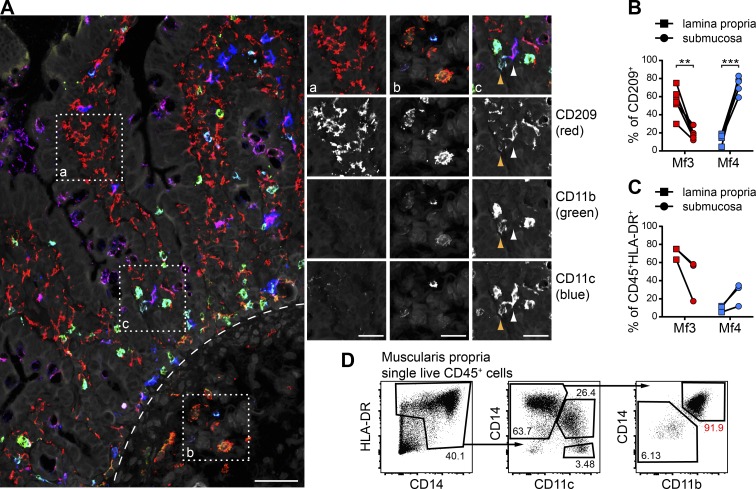
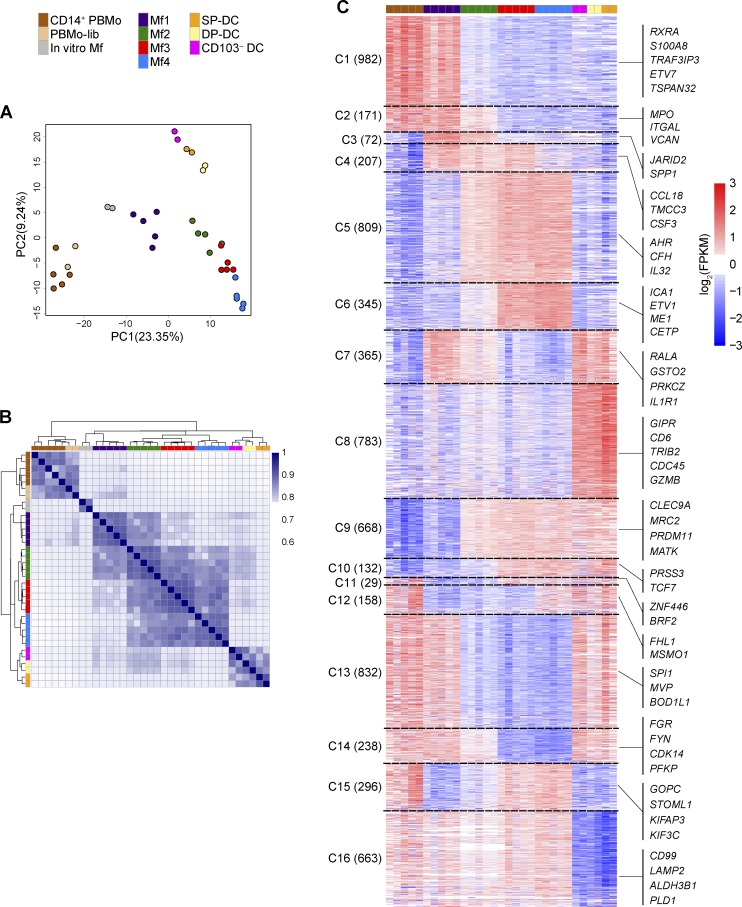
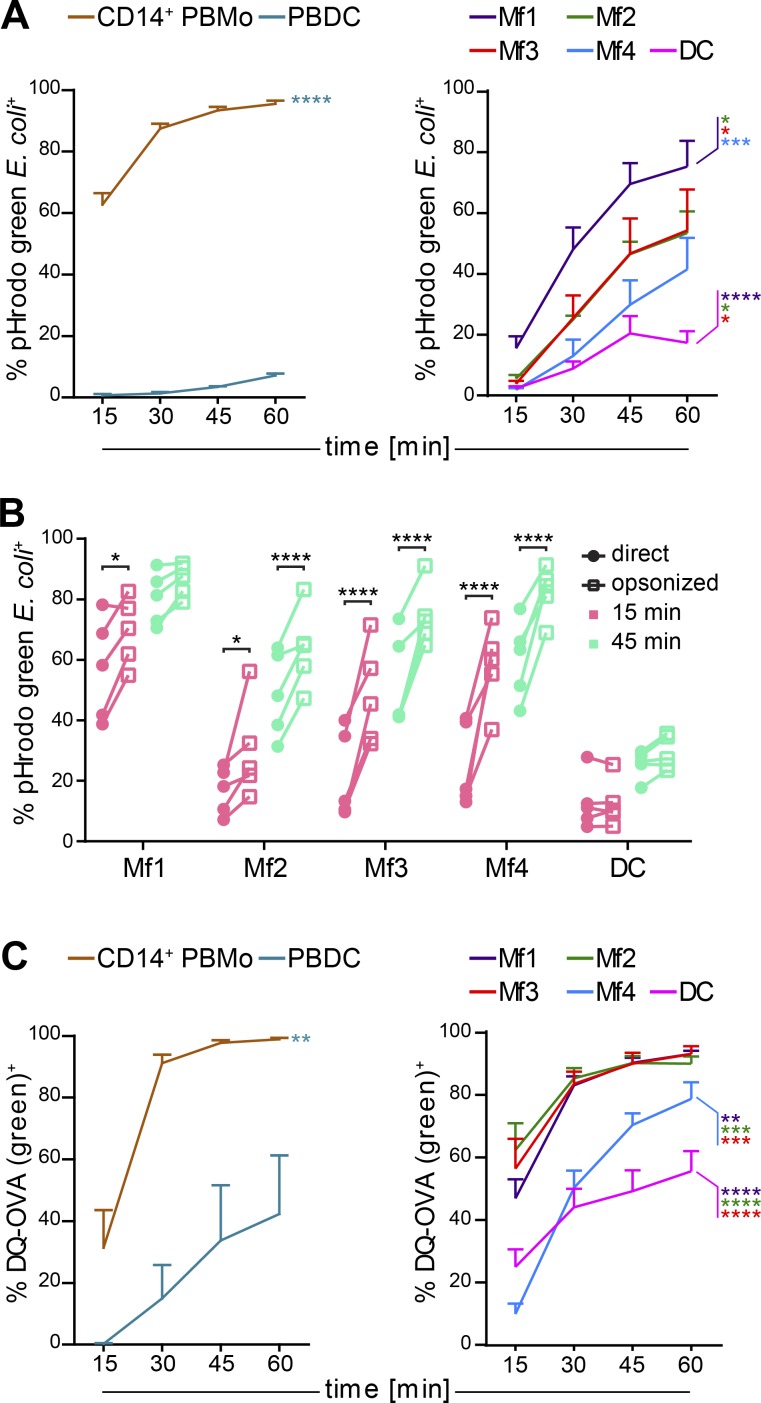
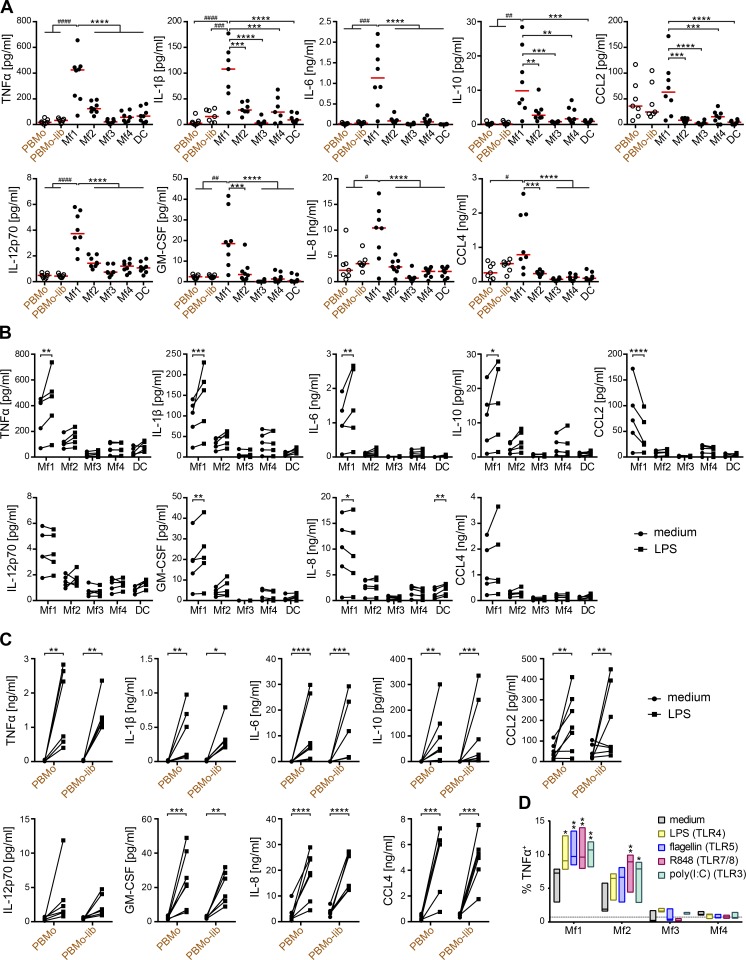
Macrophages (Mfs) are instrumental in maintaining immune homeostasis in the intestine, yet studies on the origin and heterogeneity of human intestinal Mfs are scarce. Here, we identified four distinct Mf subpopulations in human small intestine (SI). Assessment of their turnover in duodenal transplants revealed that all Mf subsets were completely replaced over time; Mf1 and Mf2, phenotypically similar to peripheral blood monocytes (PBMos), were largely replaced within 3 wk, whereas two subsets with features of mature Mfs, Mf3 and Mf4, exhibited significantly slower replacement. Mf3 and Mf4 localized differently in SI; Mf3 formed a dense network in mucosal lamina propria, whereas Mf4 was enriched in submucosa. Transcriptional analysis showed that all Mf subsets were markedly distinct from PBMos and dendritic cells. Compared with PBMos, Mf subpopulations showed reduced responsiveness to proinflammatory stimuli but were proficient at endocytosis of particulate and soluble material. These data provide a comprehensive analysis of human SI Mf population and suggest a precursor-progeny relationship with PBMos.
© 2018 Bujko et al.
Figures

References
-
- Bain C.C., Scott C.L., Uronen-Hansson H., Gudjonsson S., Jansson O., Grip O., Guilliams M., Malissen B., Agace W.W., and Mowat A.M.. 2013. Resident and pro-inflammatory macrophages in the colon represent alternative context-dependent fates of the same Ly6Chi monocyte precursors. Mucosal Immunol. 6:498–510. 10.1038/mi.2012.89 - DOI - PMC - PubMed
-
- Bain C.C., Bravo-Blas A., Scott C.L., Perdiguero E.G., Geissmann F., Henri S., Malissen B., Osborne L.C., Artis D., and Mowat A.M.. 2014. Constant replenishment from circulating monocytes maintains the macrophage pool in the intestine of adult mice. Nat. Immunol. 15:929–937. 10.1038/ni.2967 - DOI - PMC - PubMed
-
- Berg D.J., Davidson N., Kühn R., Müller W., Menon S., Holland G., Thompson-Snipes L., Leach M.W., and Rennick D.. 1996. Enterocolitis and colon cancer in interleukin-10-deficient mice are associated with aberrant cytokine production and CD4(+) TH1-like responses. J. Clin. Invest. 98:1010–1020. 10.1172/JCI118861 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
