

Morphometric Similarity Networks Detect Microscale Cortical Organization and Predict Inter-Individual Cognitive Variation
- PMID: 29276055
- PMCID: PMC5763517
- DOI: 10.1016/j.neuron.2017.11.039
Morphometric Similarity Networks Detect Microscale Cortical Organization and Predict Inter-Individual Cognitive Variation
Abstract
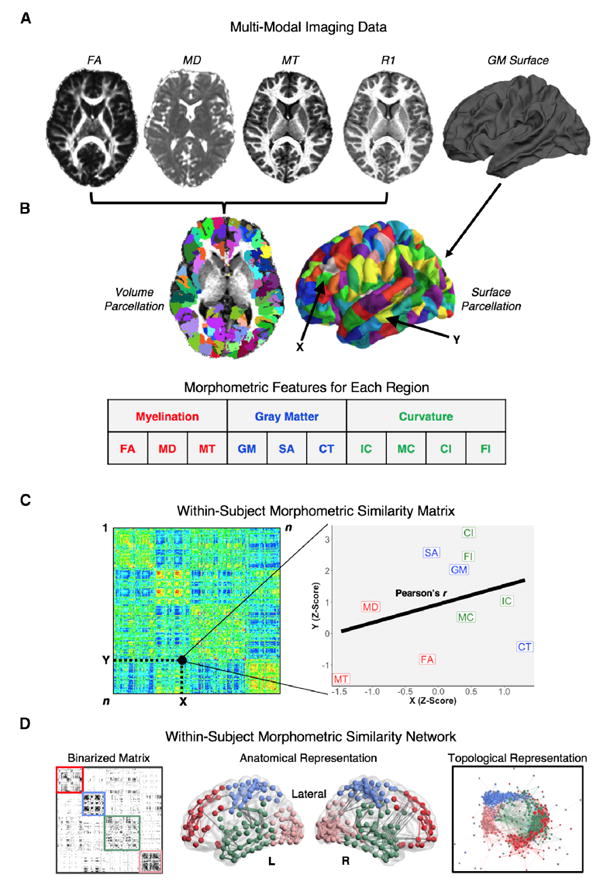
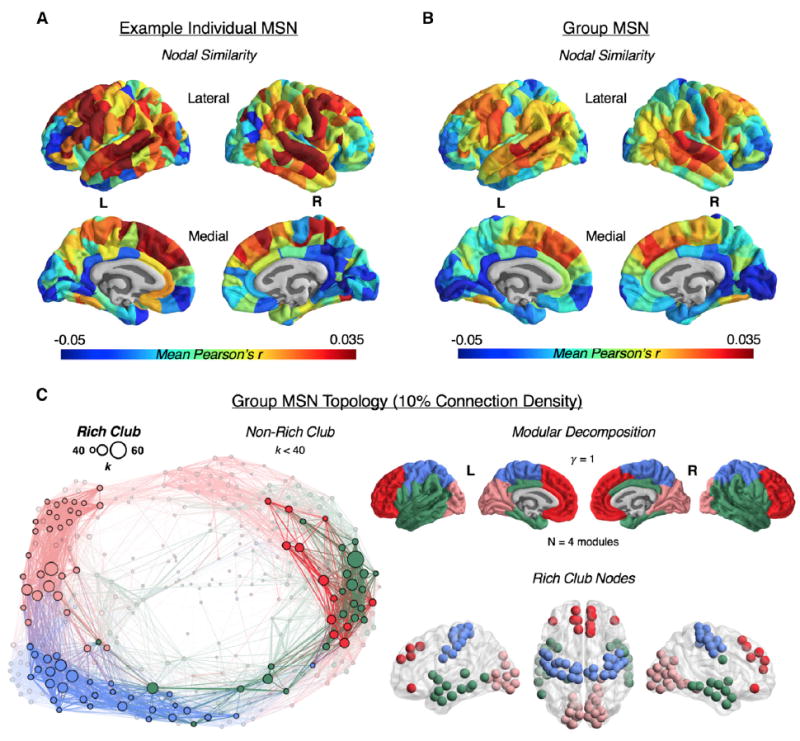
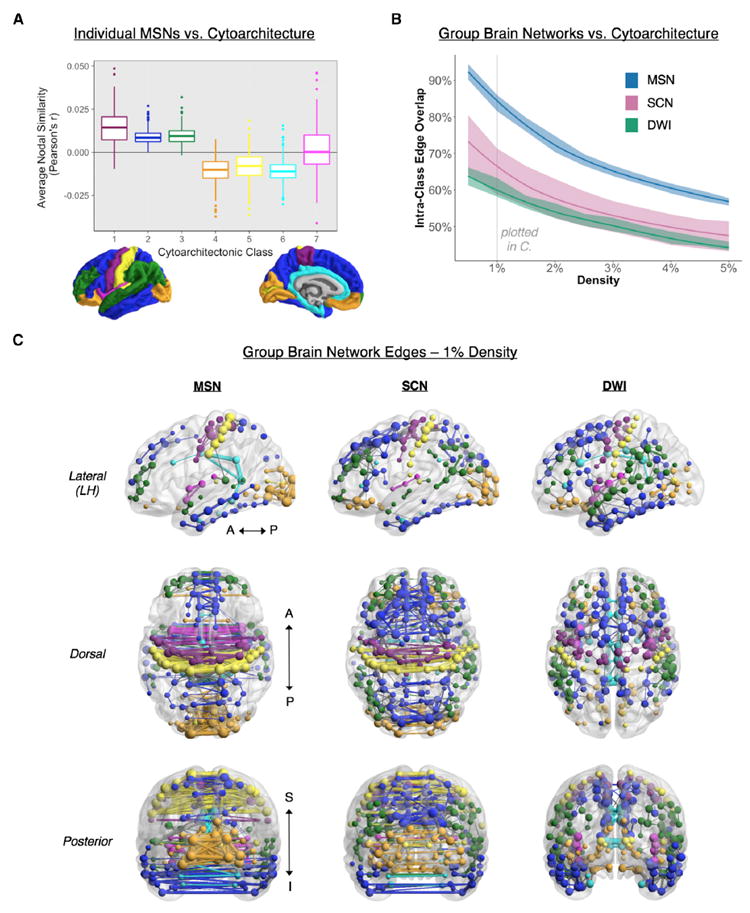
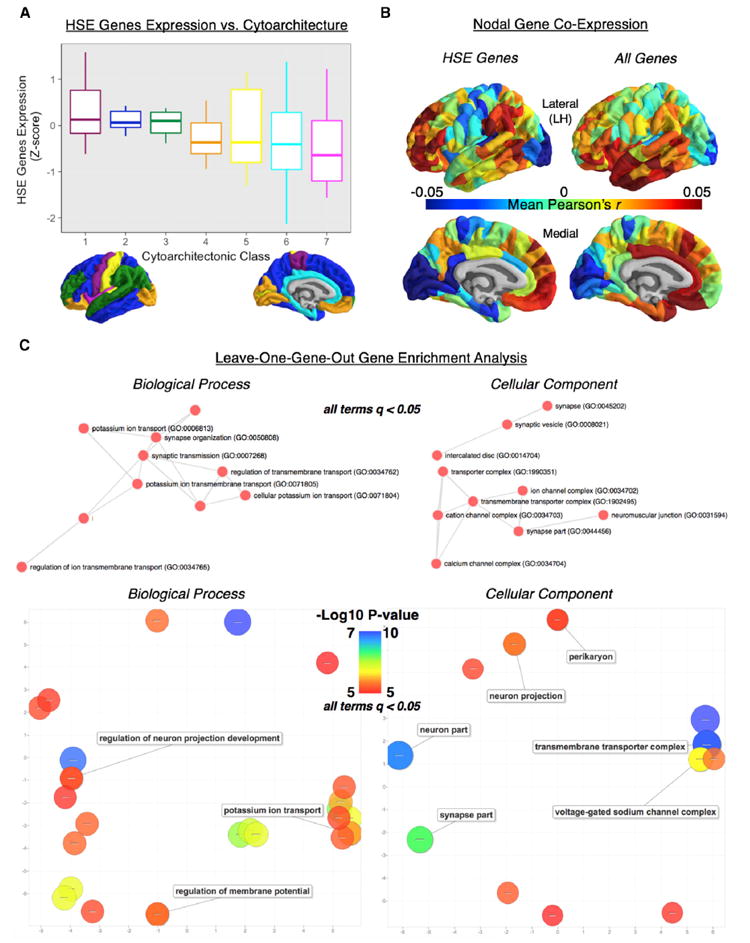
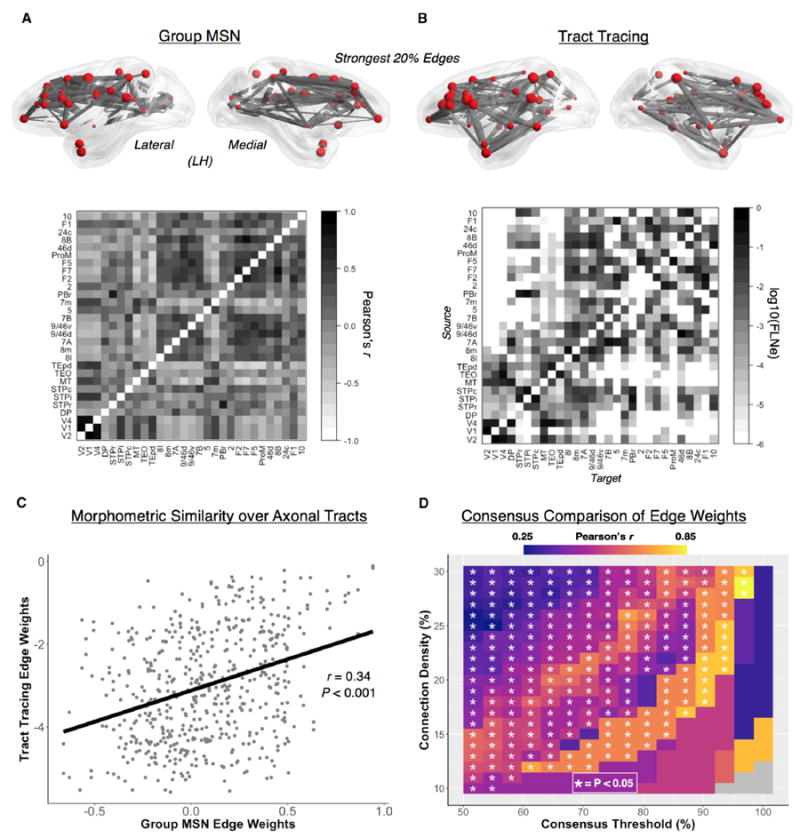
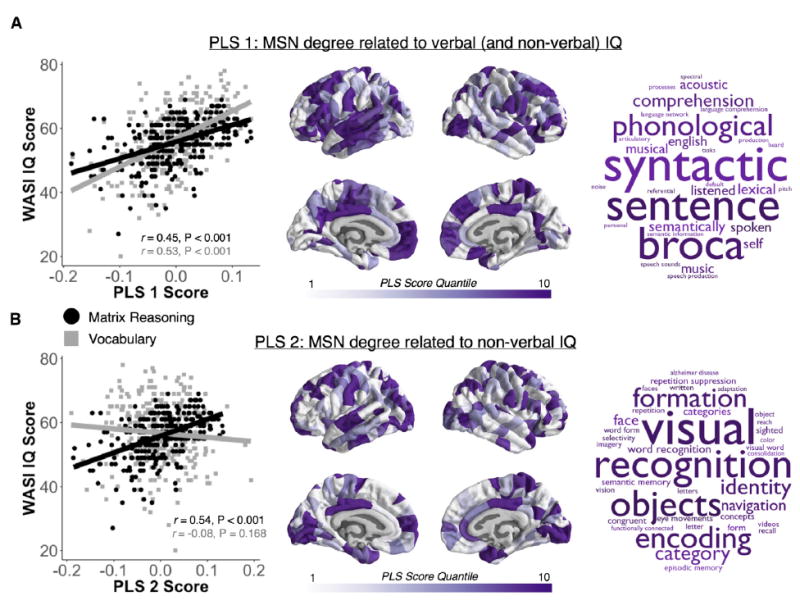
Macroscopic cortical networks are important for cognitive function, but it remains challenging to construct anatomically plausible individual structural connectomes from human neuroimaging. We introduce a new technique for cortical network mapping based on inter-regional similarity of multiple morphometric parameters measured using multimodal MRI. In three cohorts (two human, one macaque), we find that the resulting morphometric similarity networks (MSNs) have a complex topological organization comprising modules and high-degree hubs. Human MSN modules recapitulate known cortical cytoarchitectonic divisions, and greater inter-regional morphometric similarity was associated with stronger inter-regional co-expression of genes enriched for neuronal terms. Comparing macaque MSNs with tract-tracing data confirmed that morphometric similarity was related to axonal connectivity. Finally, variation in the degree of human MSN nodes accounted for about 40% of between-subject variability in IQ. Morphometric similarity mapping provides a novel, robust, and biologically plausible approach to understanding how human cortical networks underpin individual differences in psychological functions.
Keywords: IQ; MRI; connectome; cross-species; cytoarchitecture; gene expression; macaque; morphology; multi-modal.
Copyright © 2017 Elsevier Inc. All rights reserved.
Conflict of interest statement
E.T.B. is employed half-time by the University of Cambridge and half-time by GlaxoSmithKline; he holds stock in GlaxoSmithKline. I.M.G. consults to Lundbeck.
Figures
References
-
- Ad-Dab’bagh Y, Lyttelton O, Muehlboeck J, Lepage C, Einarson D, Mok K, Ivanov O, Vincent R, Lerch J, Fombonne E. In: Corbetta M, editor. The CIVET image-processing environment: a fully automated comprehensive pipeline for anatomical neuroimaging research; Proceedings of the 12th annual meeting of the Organization for Human Brain Mapping; Florence, Italy. 2006. 2266.
-
- Baars BJ. Spatial brain coherence during the establishment of a conscious event. Conscious Cogn. 1997;6:1–2. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
