

Listeria monocytogenes triggers noncanonical autophagy upon phagocytosis, but avoids subsequent growth-restricting xenophagy
- PMID: 29279409
- PMCID: PMC5777066
- DOI: 10.1073/pnas.1716055115
Listeria monocytogenes triggers noncanonical autophagy upon phagocytosis, but avoids subsequent growth-restricting xenophagy
Abstract
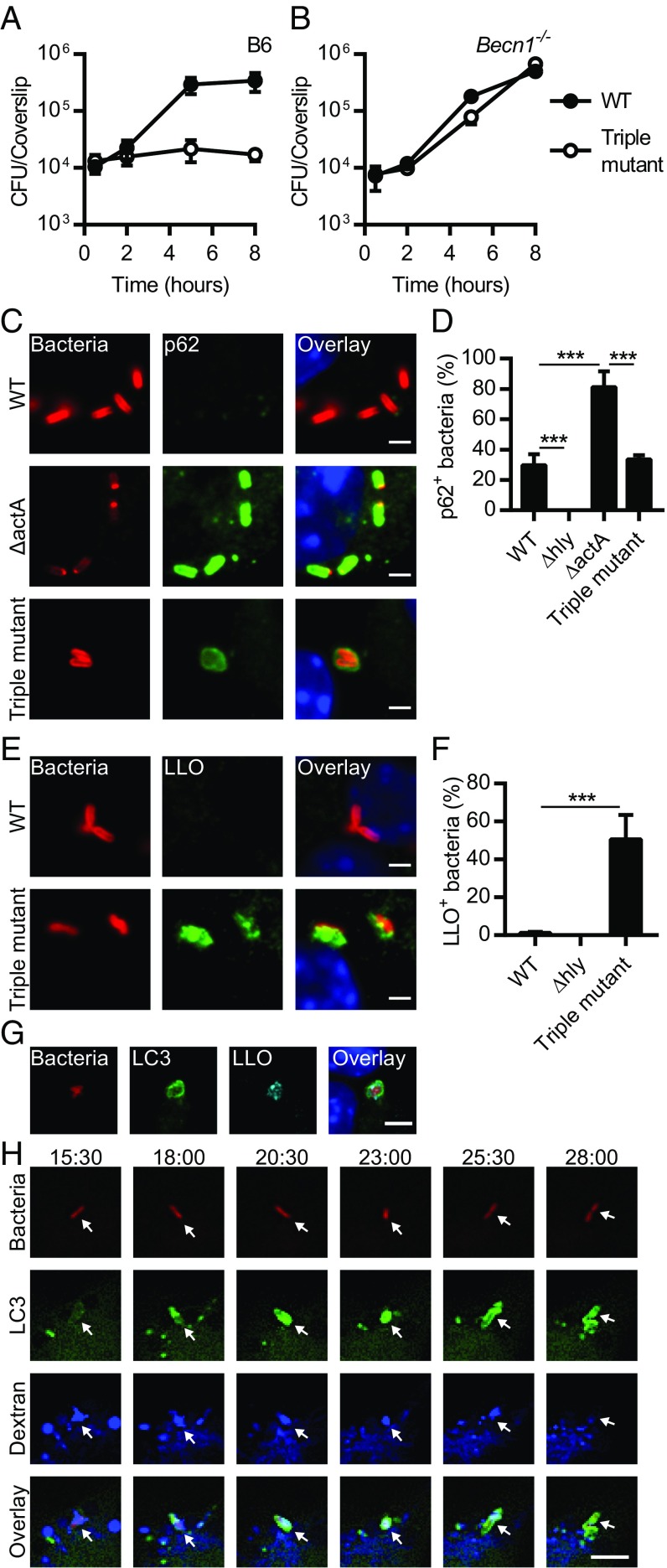
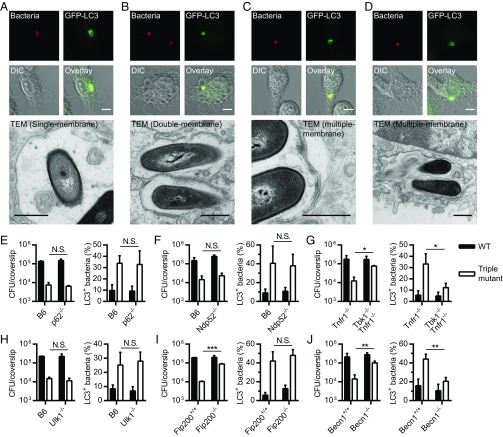
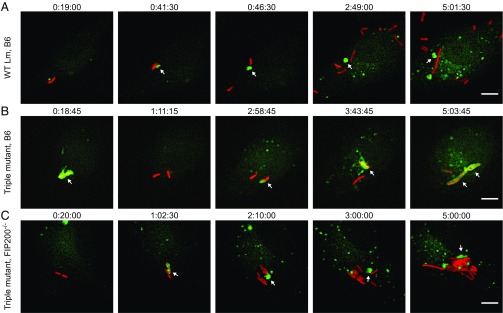
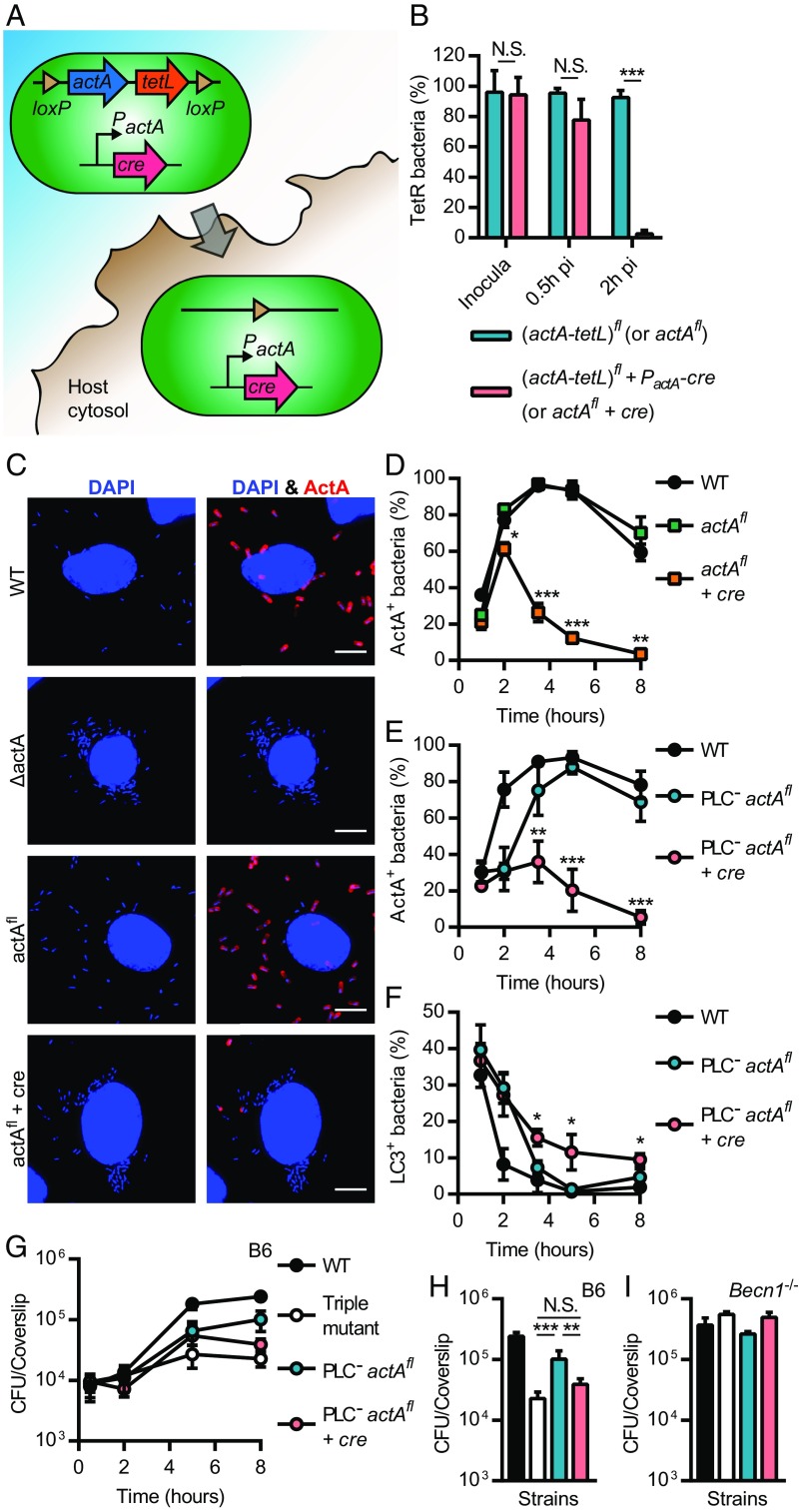
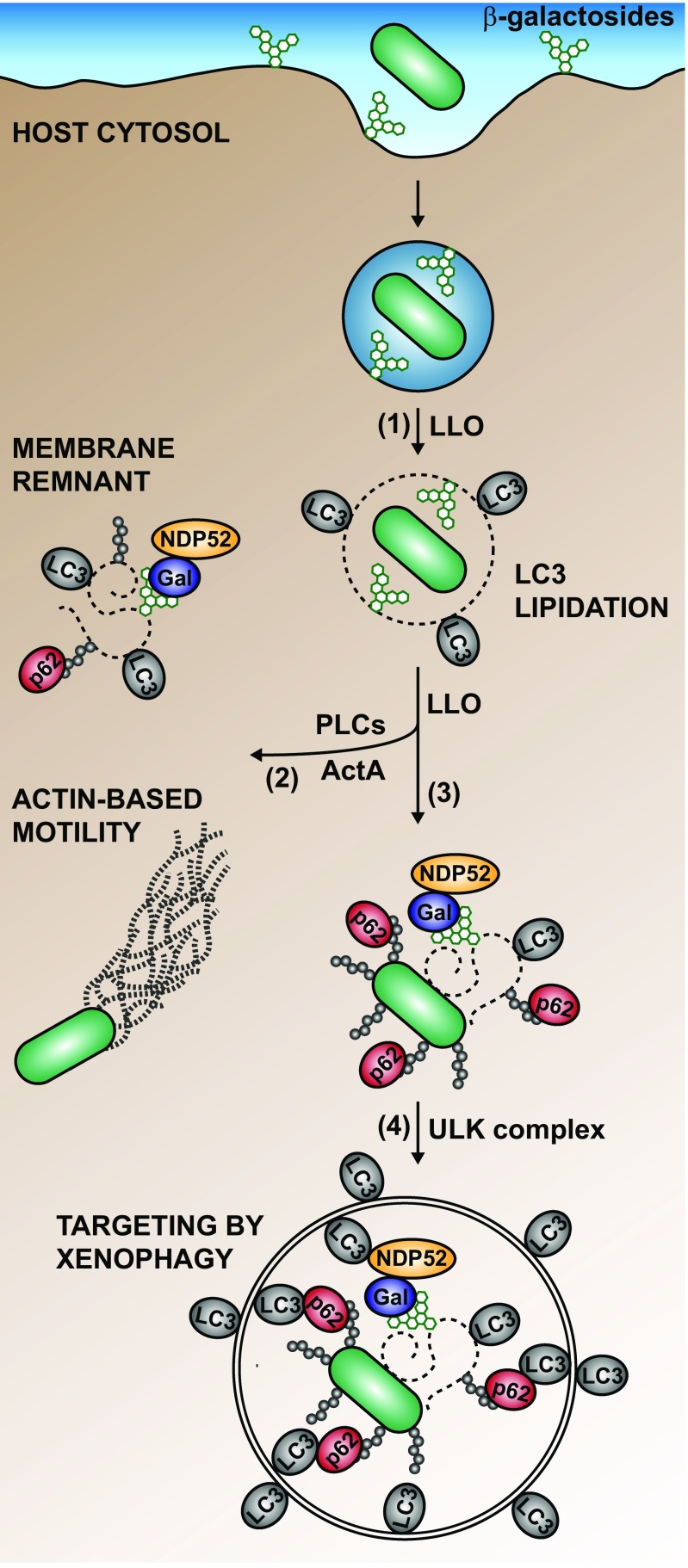
Xenophagy is a selective macroautophagic process that protects the host cytosol by entrapping and delivering microbes to a degradative compartment. Both noncanonical autophagic pathways and xenophagy are activated by microbes during infection, but the relative importance and function of these distinct processes are not clear. In this study, we used bacterial and host mutants to dissect the contribution of autophagic processes responsible for bacterial growth restriction of Listeria monocytogenesL. monocytogenes is a facultative intracellular pathogen that escapes from phagosomes, grows in the host cytosol, and avoids autophagy by expressing three determinants of pathogenesis: two secreted phospholipases C (PLCs; PlcA and PlcB) and a surface protein (ActA). We found that shortly after phagocytosis, wild-type (WT) L. monocytogenes escaped from a noncanonical autophagic process that targets damaged vacuoles. During this process, the autophagy marker LC3 localized to single-membrane phagosomes independently of the ULK complex, which is required for initiation of macroautophagy. However, growth restriction of bacteria lacking PlcA, PlcB, and ActA required FIP200 and TBK1, both involved in the engulfment of microbes by xenophagy. Time-lapse video microscopy revealed that deposition of LC3 on L. monocytogenes-containing vacuoles via noncanonical autophagy had no apparent role in restricting bacterial growth and that, upon access to the host cytosol, WT L. monocytogenes utilized PLCs and ActA to avoid subsequent xenophagy. In conclusion, although noncanonical autophagy targets phagosomes, xenophagy was required to restrict the growth of L. monocytogenes, an intracellular pathogen that damages the entry vacuole.
Keywords: ActA; LC3-associated phagocytosis; bacteria; macrophage; phospholipases.
Conflict of interest statement
Conflict of interest statement: D.A.P. has a consulting relationship with and a financial interest in Aduro Biotech. Both he and the company stand to benefit from the commercialization of the results of this research. G.M. and Ralph R. Isberg are coauthors on a 2017 review article.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
