

ALKBH5-dependent m6A demethylation controls splicing and stability of long 3'-UTR mRNAs in male germ cells
- PMID: 29279410
- PMCID: PMC5777073
- DOI: 10.1073/pnas.1717794115
ALKBH5-dependent m6A demethylation controls splicing and stability of long 3'-UTR mRNAs in male germ cells
Abstract
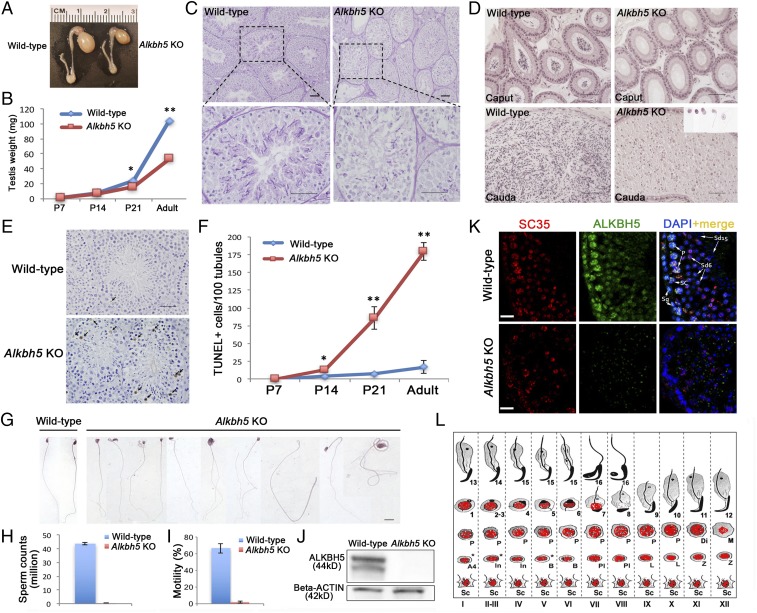
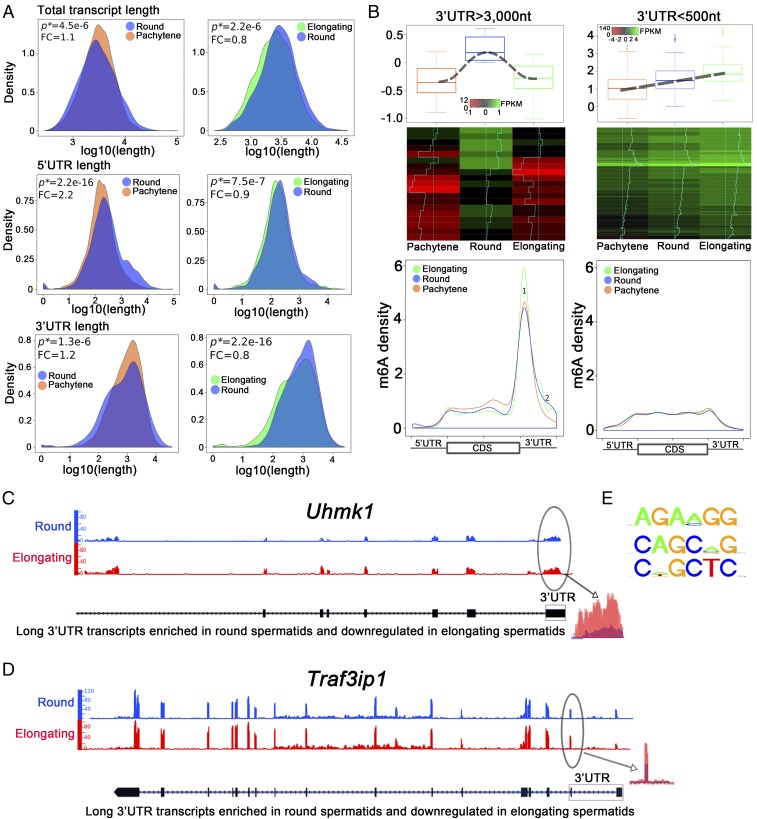
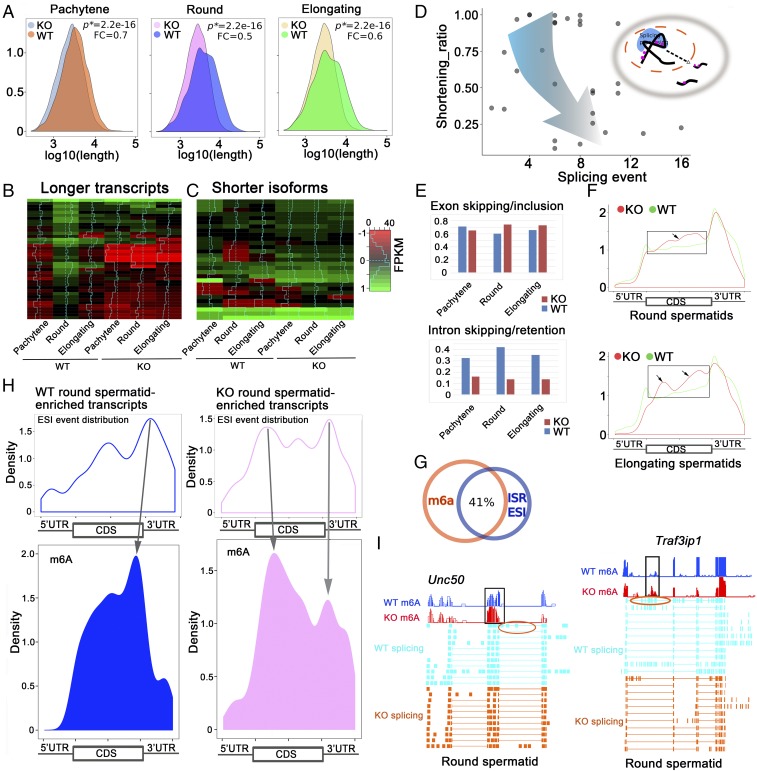
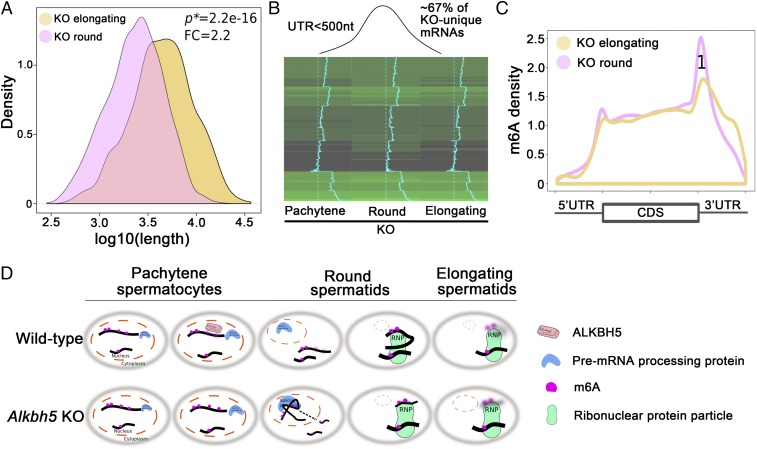
N6-methyladenosine (m6A) represents one of the most common RNA modifications in eukaryotes. Specific m6A writer, eraser, and reader proteins have been identified. As an m6A eraser, ALKBH5 specifically removes m6A from target mRNAs and inactivation of Alkbh5 leads to male infertility in mice. However, the underlying molecular mechanism remains unknown. Here, we report that ALKBH5-mediated m6A erasure in the nuclei of spermatocytes and round spermatids is essential for correct splicing and the production of longer 3'-UTR mRNAs, and failure to do so leads to aberrant splicing and production of shorter transcripts with elevated levels of m6A that are rapidly degraded. Our study identified reversible m6A modification as a critical mechanism of posttranscriptional control of mRNA fate in late meiotic and haploid spermatogenic cells.
Keywords: 3′-UTR shortening; RNA methylation; alternative splicing; fertility; mRNA stability.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Overexpression of m6A-factors METTL3, ALKBH5, and YTHDC1 alters HPV16 mRNA splicing.Virus Genes. 2022 Apr;58(2):98-112. doi: 10.1007/s11262-022-01889-6. Epub 2022 Feb 21. Virus Genes. 2022. PMID: 35190939 Free PMC article.
-
RNA demethylase ALKBH5 prevents pancreatic cancer progression by posttranscriptional activation of PER1 in an m6A-YTHDF2-dependent manner.Mol Cancer. 2020 May 19;19(1):91. doi: 10.1186/s12943-020-01158-w. Mol Cancer. 2020. PMID: 32429928 Free PMC article.
-
m6A modifications regulate intestinal immunity and rotavirus infection.Elife. 2022 Jan 31;11:e73628. doi: 10.7554/eLife.73628. Elife. 2022. PMID: 35098923 Free PMC article.
-
Role of the m6A demethylase ALKBH5 in gastrointestinal tract cancer (Review).Int J Mol Med. 2025 Feb;55(2):22. doi: 10.3892/ijmm.2024.5463. Epub 2024 Nov 29. Int J Mol Med. 2025. PMID: 39611478 Free PMC article. Review.
-
RNA demethylase ALKBH5 in cancer: from mechanisms to therapeutic potential.J Hematol Oncol. 2022 Jan 21;15(1):8. doi: 10.1186/s13045-022-01224-4. J Hematol Oncol. 2022. PMID: 35063010 Free PMC article. Review.
Cited by
-
The Role of RNA Methyltransferase METTL3 in Hepatocellular Carcinoma: Results and Perspectives.Front Cell Dev Biol. 2021 May 11;9:674919. doi: 10.3389/fcell.2021.674919. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34046411 Free PMC article. Review.
-
Demethyltransferase AlkBH1 substrate diversity and relationship to human diseases.Mol Biol Rep. 2021 May;48(5):4747-4756. doi: 10.1007/s11033-021-06421-x. Epub 2021 May 27. Mol Biol Rep. 2021. PMID: 34046849 Review.
-
Modeling multi-species RNA modification through multi-task curriculum learning.Nucleic Acids Res. 2021 Apr 19;49(7):3719-3734. doi: 10.1093/nar/gkab124. Nucleic Acids Res. 2021. PMID: 33744973 Free PMC article.
-
Dealkylation of Macromolecules by Eukaryotic α-Ketoglutarate-Dependent Dioxygenases from the AlkB-like Family.Curr Issues Mol Biol. 2024 Sep 20;46(9):10462-10491. doi: 10.3390/cimb46090622. Curr Issues Mol Biol. 2024. PMID: 39329974 Free PMC article. Review.
-
Epigenetic regulation of cancer stem cell and tumorigenesis.Adv Cancer Res. 2020;148:1-26. doi: 10.1016/bs.acr.2020.05.001. Epub 2020 Jul 16. Adv Cancer Res. 2020. PMID: 32723561 Free PMC article. Review.
References
-
- He C. Grand challenge commentary: RNA epigenetics? Nat Chem Biol. 2010;6:863–865. - PubMed
-
- Rottman F, Shatkin AJ, Perry RP. Sequences containing methylated nucleotides at the 5′ termini of messenger RNAs: Possible implications for processing. Cell. 1974;3:197–199. - PubMed
-
- Wei CM, Gershowitz A, Moss B. Methylated nucleotides block 5′ terminus of HeLa cell messenger RNA. Cell. 1975;4:379–386. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
