

Subtle variation in shade avoidance responses may have profound consequences for plant competitiveness
- PMID: 29280992
- PMCID: PMC5906909
- DOI: 10.1093/aob/mcx151
Subtle variation in shade avoidance responses may have profound consequences for plant competitiveness
Abstract
Background and aims: Although phenotypic plasticity has been shown to be beneficial for plant competitiveness for light, there is limited knowledge on how variation in these plastic responses plays a role in determining competitiveness.
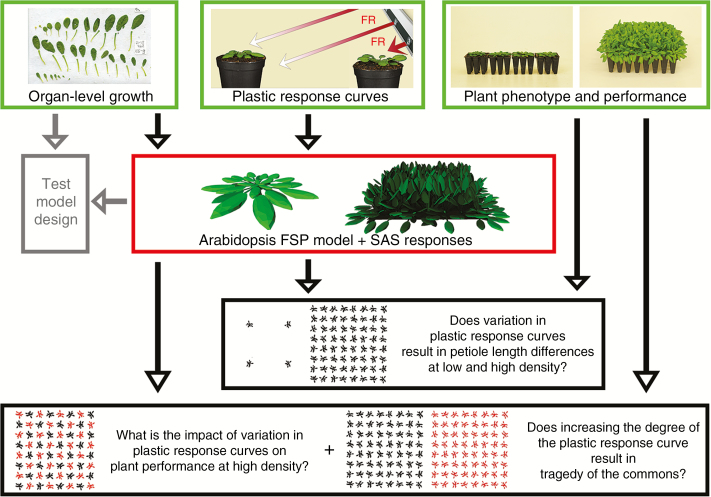
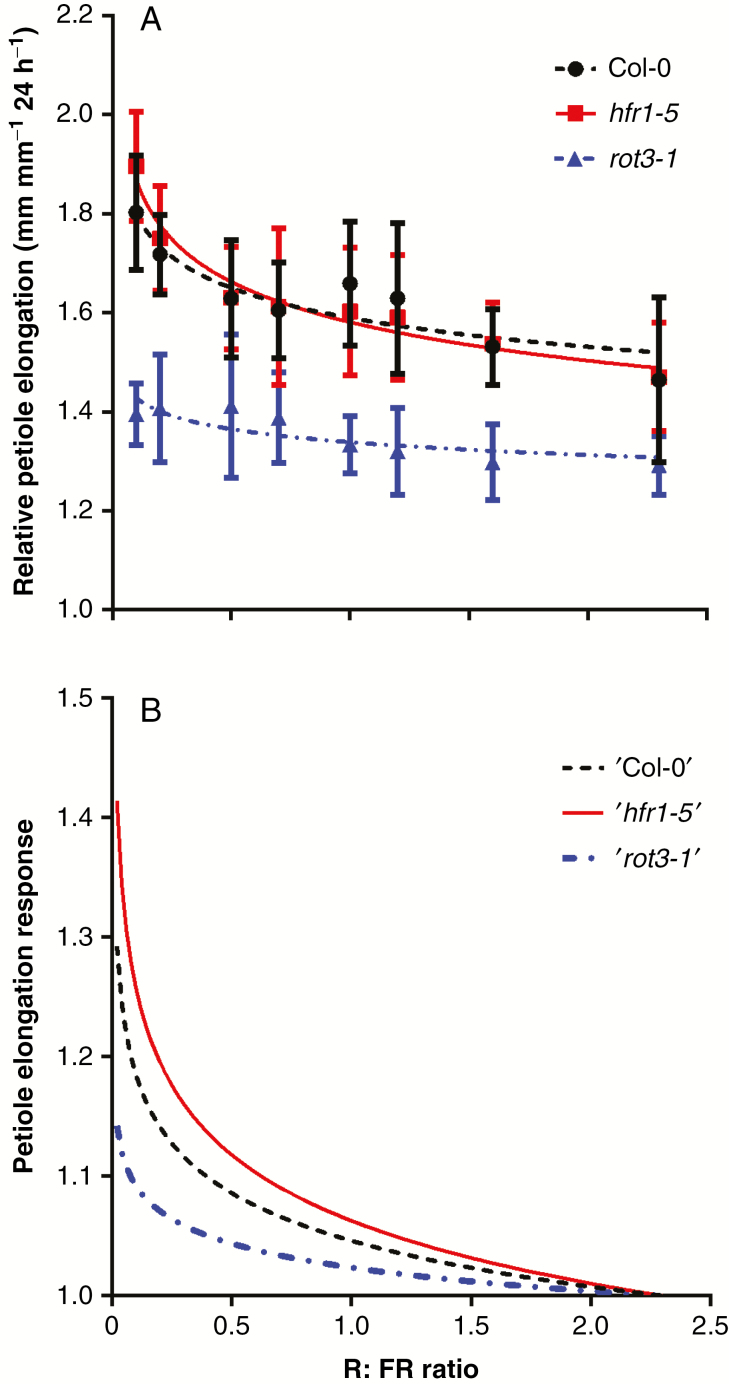
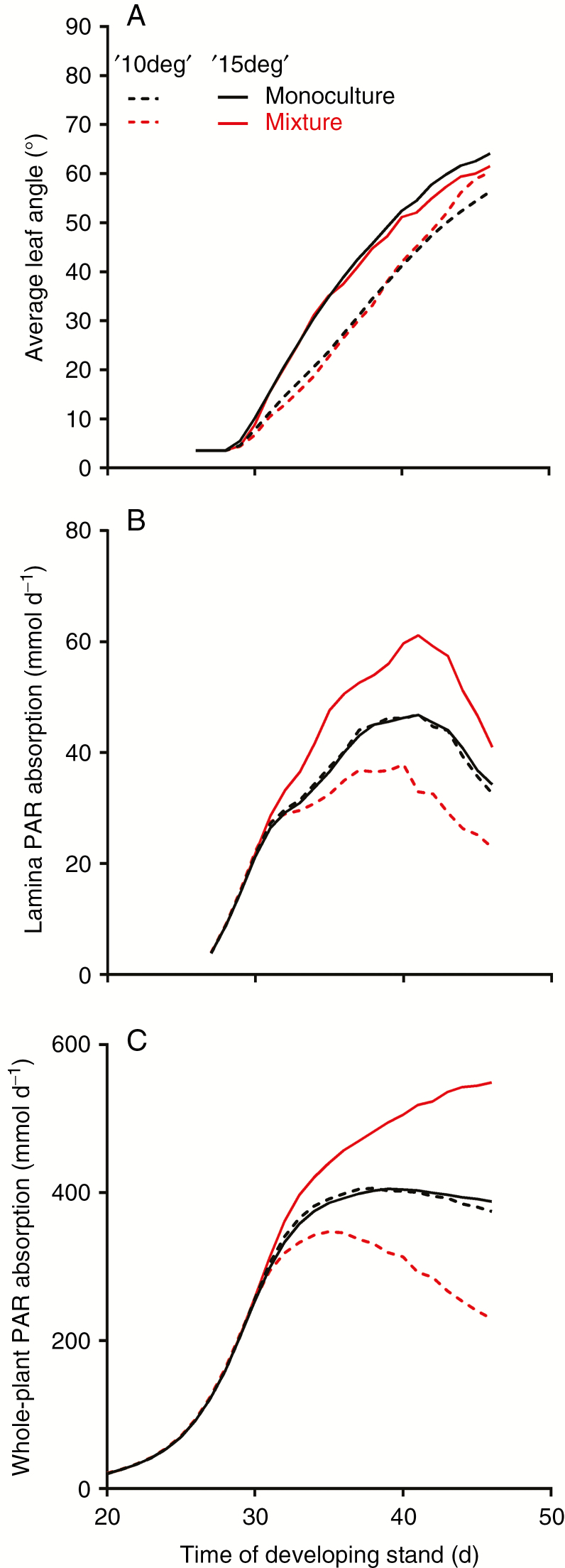
Methods: A combination of detailed plant experiments and functional-structural plant (FSP) modelling was used that captures the complex dynamic feedback between the changing plant phenotype and the within-canopy light environment in time and 3-D space. Leaf angle increase (hyponasty) and changes in petiole elongation rates in response to changes in the ratio between red and far-red light, two important shade avoidance responses in Arabidopsis thaliana growing in dense population stands, were chosen as a case study for plant plasticity. Measuring and implementing these responses into an FSP model allowed simulation of plant phenotype as an emergent property of the underlying growth and response mechanisms.
Key results: Both the experimental and model results showed that substantial differences in competitiveness may arise between genotypes with only marginally different hyponasty or petiole elongation responses, due to the amplification of plant growth differences by small changes in plant phenotype. In addition, this study illustrated that strong competitive responses do not necessarily have to result in a tragedy of the commons; success in competition at the expense of community performance.
Conclusions: Together, these findings indicate that selection pressure could probably have played a role in fine-tuning the sensitive shade avoidance responses found in plants. The model approach presented here provides a novel tool to analyse further how natural selection could have acted on the evolution of plastic responses.
Figures





References
-
- Anten NPR, Vermeulen PJ. 2016. Tragedies and crops: understanding natural selection to improve cropping systems. Trends in Ecology and Evolution 31: 429–439. - PubMed
-
- Anten NPR, Schieving F, Werger MJA. 1995. Patterns of light and nitrogen distribution in relation to whole canopy carbon gain in C3 and C4 mono- and dicotyledonous species. Oecologia 101: 504–513. - PubMed
-
- Ballaré CL, Pierik R. 2017. The shade-avoidance syndrome: multiple signals and ecological outputs. Plant, Cell and Environment 40: 2530–2543. - PubMed
-
- Bardgett RD, Mommer L, De Vries FT. 2014. Going underground: root traits as drivers of ecosystem processes. Trends in Ecology and Evolution 29: 692–699. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
