

Transcriptomic and Metabolomic Reprogramming from Roots to Haustoria in the Parasitic Plant, Thesium chinense
- PMID: 29281058
- PMCID: PMC6018956
- DOI: 10.1093/pcp/pcx200
Transcriptomic and Metabolomic Reprogramming from Roots to Haustoria in the Parasitic Plant, Thesium chinense
Erratum in
-
Erratum.Plant Cell Physiol. 2018 Apr 1;59(4):876. doi: 10.1093/pcp/pcy075. Plant Cell Physiol. 2018. PMID: 29718476 Free PMC article. No abstract available.
Abstract
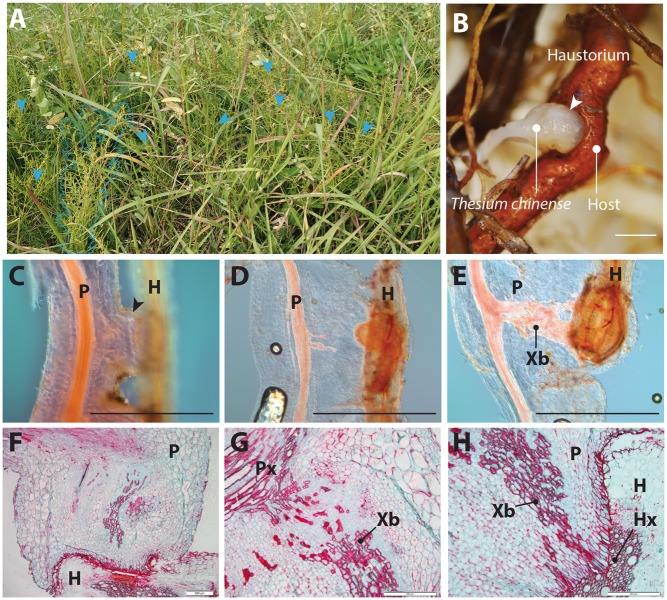
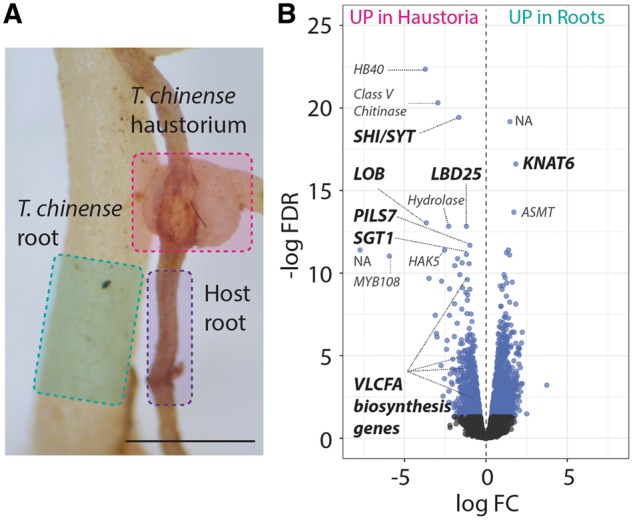
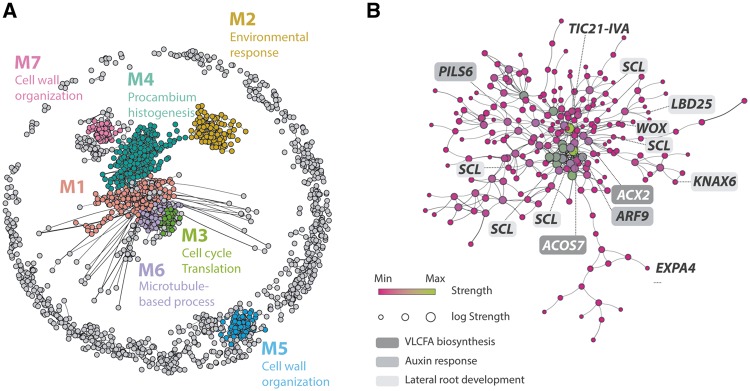
Most plants show remarkable developmental plasticity in the generation of diverse types of new organs upon external stimuli, allowing them to adapt to their environment. Haustorial formation in parasitic plants is an example of such developmental reprogramming, but its molecular mechanism is largely unknown. In this study, we performed field-omics using transcriptomics and metabolomics to profile the molecular switch occurring in haustorial formation of the root parasitic plant, Thesium chinense, collected from its natural habitat. RNA-sequencing with de novo assembly revealed that the transcripts of very long chain fatty acid (VLCFA) biosynthesis genes, auxin biosynthesis/signaling-related genes and lateral root developmental genes are highly abundant in the haustoria. Gene co-expression network analysis identified a network module linking VLCFAs and the auxin-responsive lateral root development pathway. GC-TOF-MS analysis consistently revealed a unique metabolome profile with many types of fatty acids in the T. chinense root system, including the accumulation of a 25-carbon long chain saturated fatty acid in the haustoria. Our field-omics data provide evidence supporting the hypothesis that the molecular developmental machinery used for lateral root formation in non-parasitic plants has been co-opted into the developmental reprogramming of haustorial formation in the linage of parasitic plants.
Figures

References
-
- Aly R., Cholakh H., Joel D.M., Leibman D., Steinitz B., Zelcer A., et al. (2009) Gene silencing of mannose 6-phosphate reductase in the parasitic weed Orobanche aegyptiaca through the production of homologous dsRNA sequences in the host plant. Plant Biotechnol. J. 7: 487–498. - PubMed
-
- Baldovini N., Delasalle C., Joulain D. (2011) Phytochemistry of the heartwood from fragrant Santalum species: a review. Flavour Fragr. J. 26: 7–26.
-
- Bandaranayake P.C., Yoder J.I. (2013) Trans-specific gene silencing of acetyl-CoA carboxylase in a root-parasitic plant. Mol. Plant Microbe Interact. 26: 575–584. - PubMed
-
- Barbez E., Kubes M., Rolcik J., Beziat C., Pencik A., Wang B., et al. (2012) A novel putative auxin carrier family regulates intracellular auxin homeostasis in plants. Nature 485: 119–122. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
