

Identification of Oxa1 Homologs Operating in the Eukaryotic Endoplasmic Reticulum
- PMID: 29281821
- PMCID: PMC5868721
- DOI: 10.1016/j.celrep.2017.12.006
Identification of Oxa1 Homologs Operating in the Eukaryotic Endoplasmic Reticulum
Abstract
Members of the evolutionarily conserved Oxa1/Alb3/YidC family mediate membrane protein biogenesis at the mitochondrial inner membrane, chloroplast thylakoid membrane, and bacterial plasma membrane, respectively. Despite their broad phylogenetic distribution, no Oxa1/Alb3/YidC homologs are known to operate in eukaryotic cells outside the endosymbiotic organelles. Here, we present bioinformatic evidence that the tail-anchored protein insertion factor WRB/Get1, the "endoplasmic reticulum (ER) membrane complex" subunit EMC3, and TMCO1 are ER-resident homologs of the Oxa1/Alb3/YidC family. Topology mapping and co-evolution-based modeling demonstrate that Get1, EMC3, and TMCO1 share a conserved Oxa1-like architecture. Biochemical analysis of human TMCO1, the only homolog not previously linked to membrane protein biogenesis, shows that it associates with the Sec translocon and ribosomes. These findings suggest a specific biochemical function for TMCO1 and define a superfamily of proteins-the "Oxa1 superfamily"-whose shared function is to facilitate membrane protein biogenesis.
Copyright © 2017 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
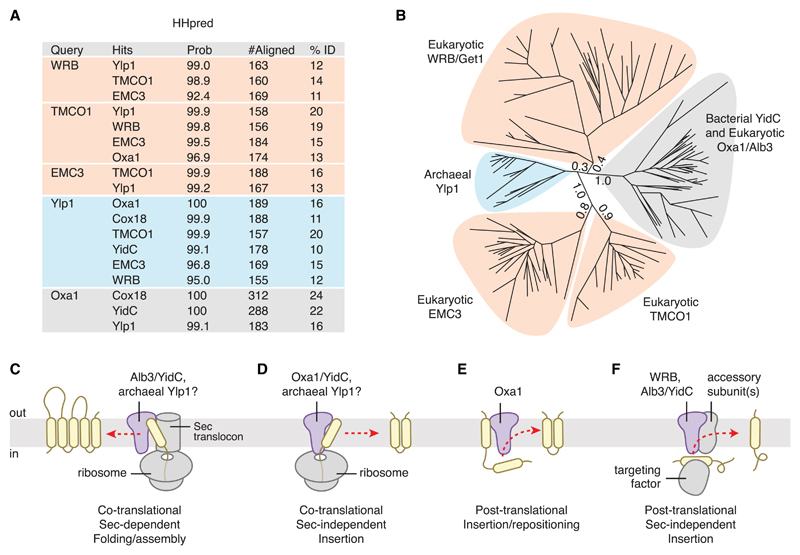
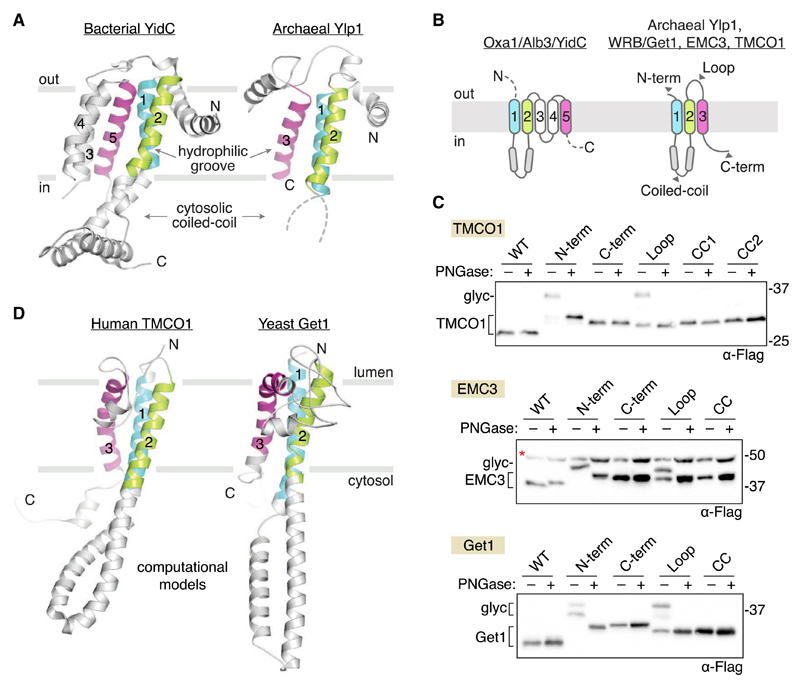
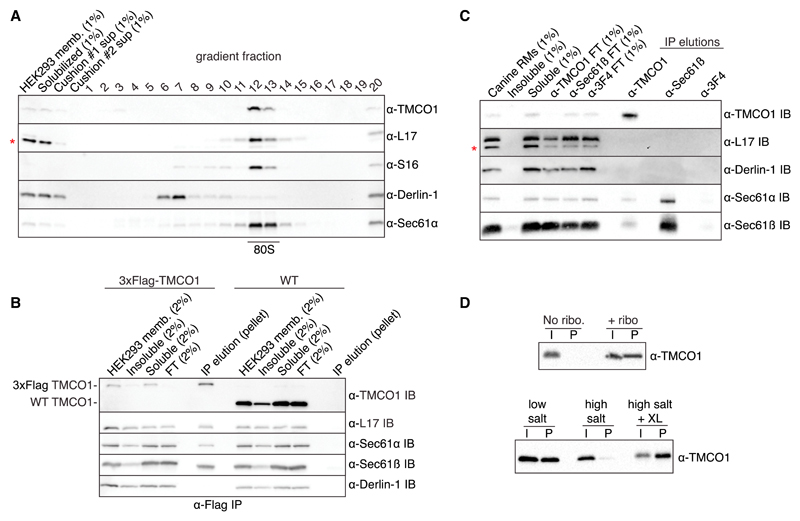
Comment in
-
Oxa1 Superfamily: New Members Found in the ER.Trends Biochem Sci. 2018 Mar;43(3):151-153. doi: 10.1016/j.tibs.2017.12.005. Epub 2018 Jan 12. Trends Biochem Sci. 2018. PMID: 29310909
References
-
- Alanay Y, Ergüner B, Utine E, Haçarız O, Kiper POS, Taşkıran EZ, Perçin F, Uz E, Sağıroğlu MŞ, Yuksel B, et al. TMCO1 deficiency causes autosomal recessive cerebrofaciothoracic dysplasia. Am J Med Genet A. 2014;164:291–304. - PubMed
-
- Aviram N, Schuldiner M. Embracing the void–how much do we really know about targeting and translocation to the endoplasmic reticulum? Curr Opin Cell Biol. 2014;29:8–17. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
