

Molecular characterization of perivascular drainage pathways in the murine brain
- PMID: 29283289
- PMCID: PMC5888861
- DOI: 10.1177/0271678X17749689
Molecular characterization of perivascular drainage pathways in the murine brain
Erratum in
-
Erratum.J Cereb Blood Flow Metab. 2018 Aug;38(8):1384. doi: 10.1177/0271678X18775331. J Cereb Blood Flow Metab. 2018. PMID: 30066612 Free PMC article.
Abstract
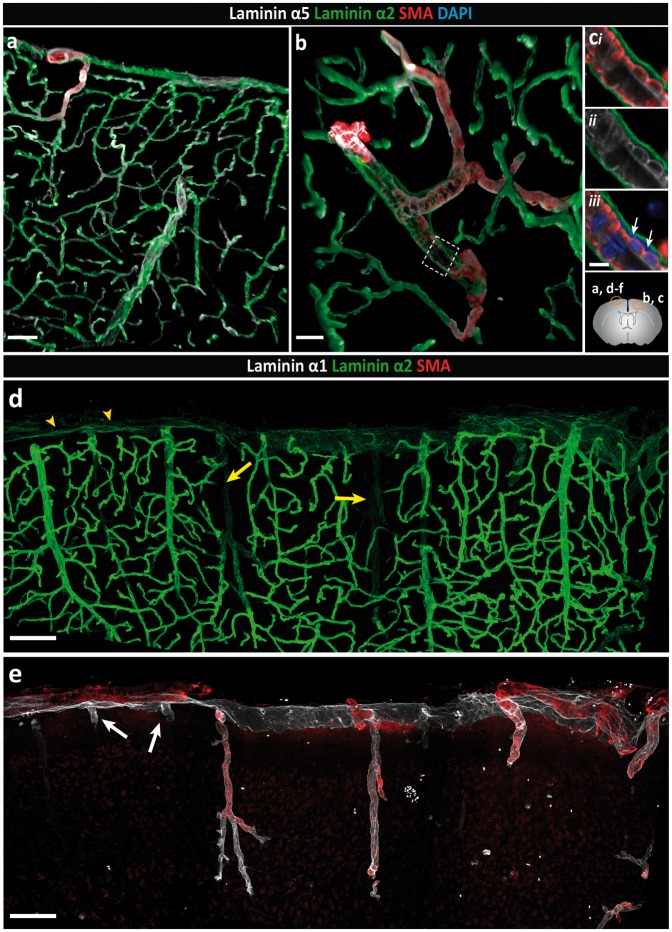
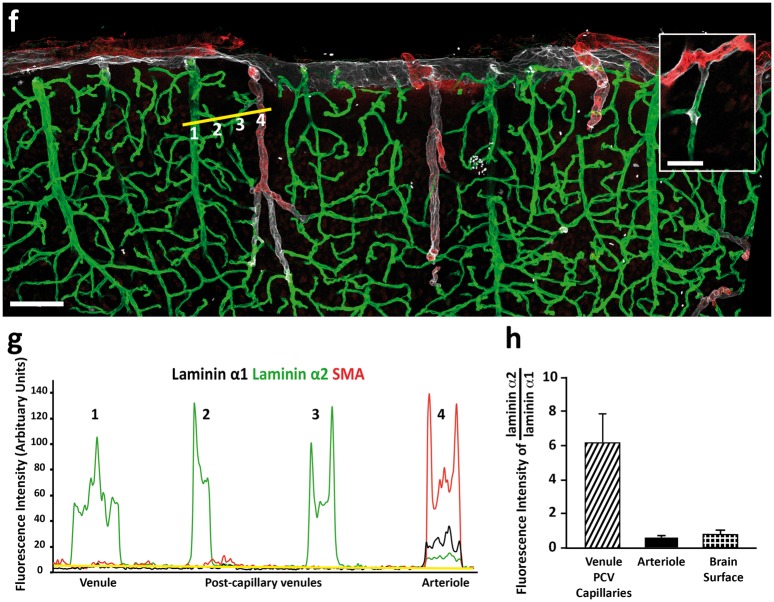
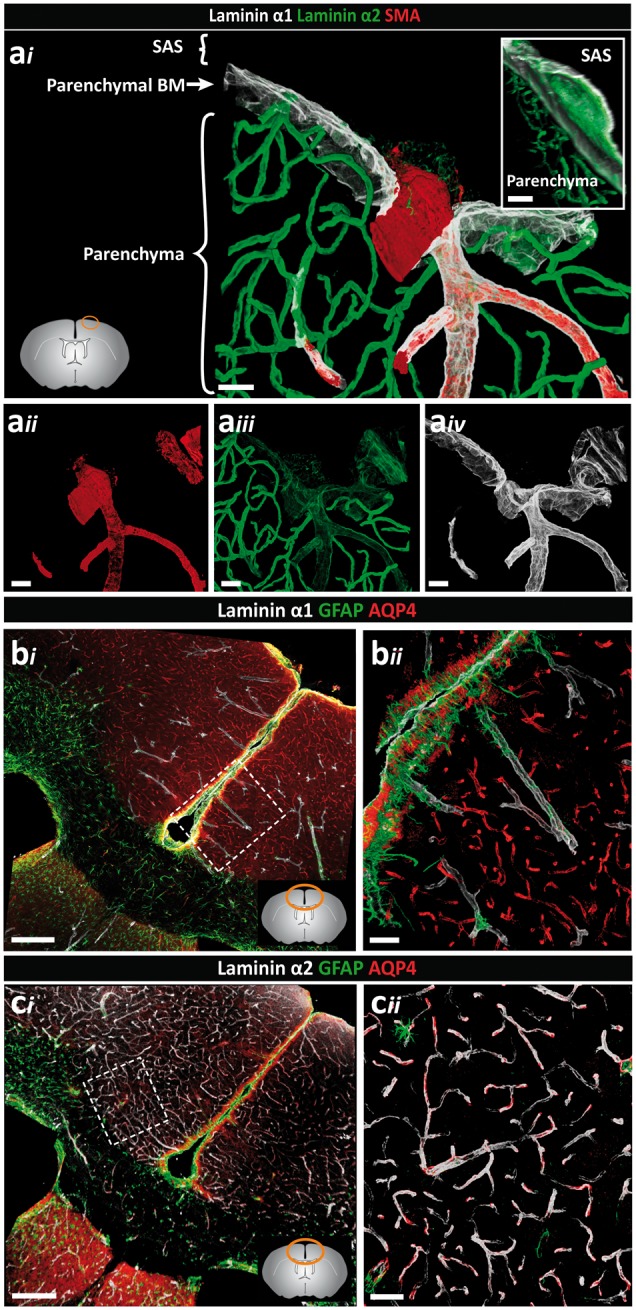
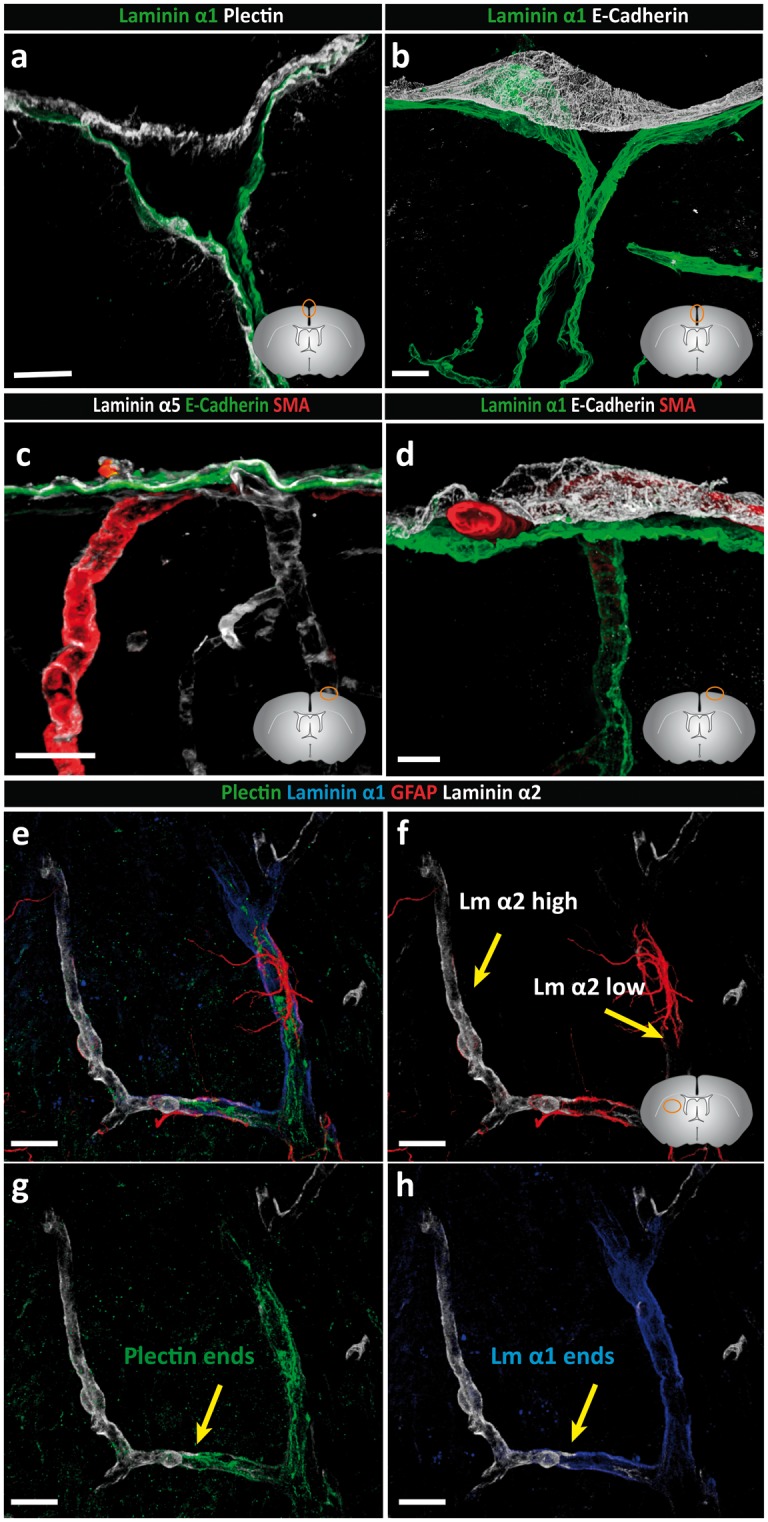
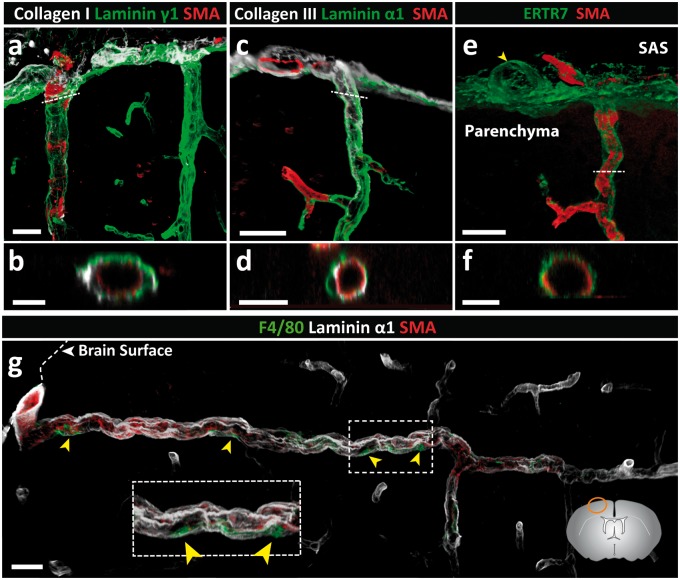
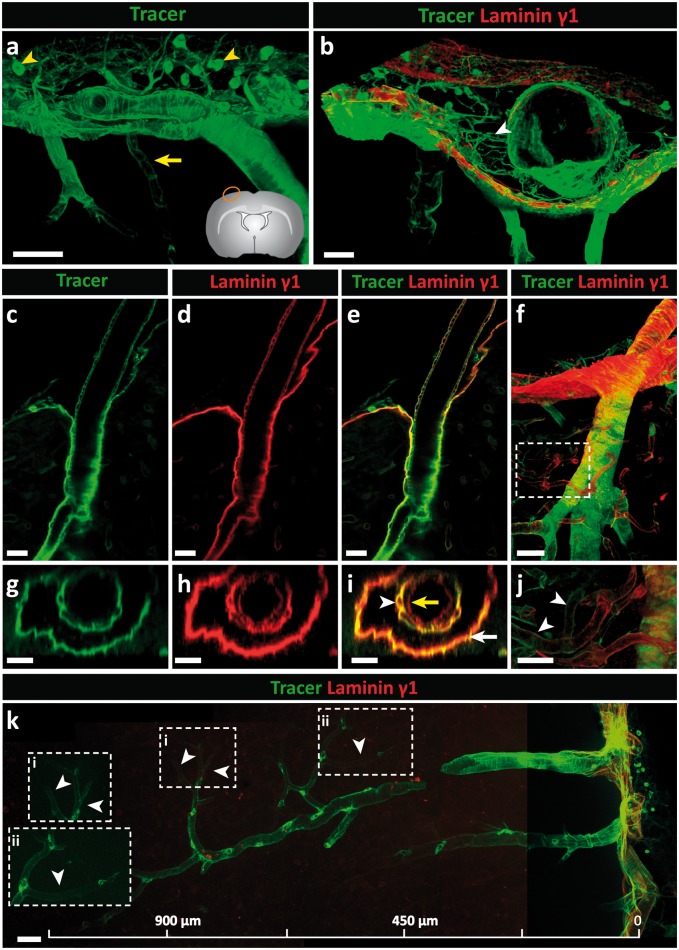
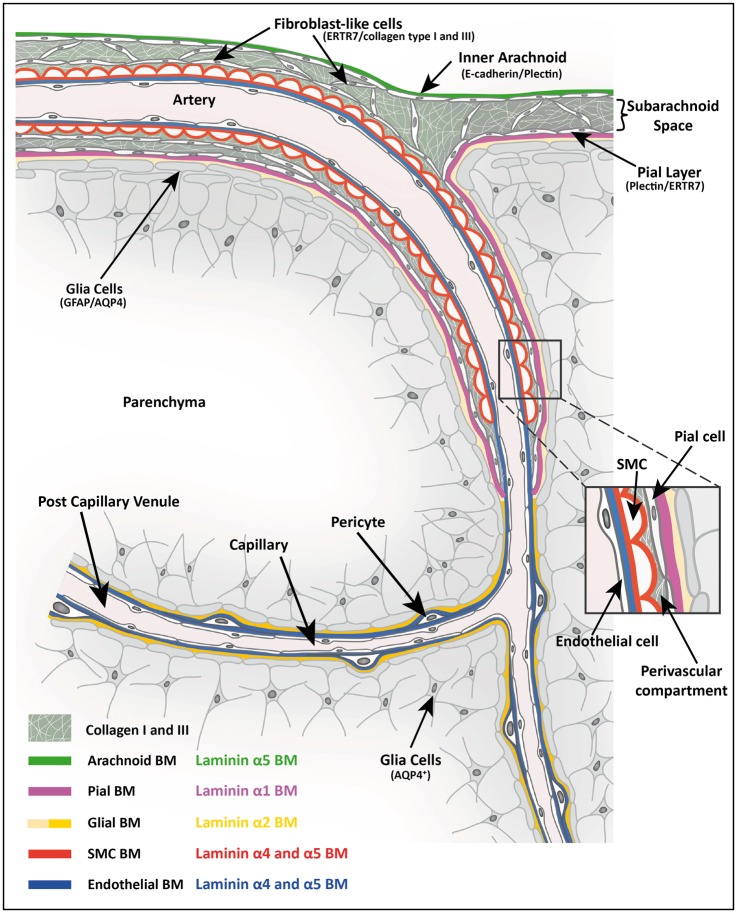
Perivascular compartments surrounding central nervous system (CNS) vessels have been proposed to serve key roles in facilitating cerebrospinal fluid flow into the brain, CNS waste transfer, and immune cell trafficking. Traditionally, these compartments were identified by electron microscopy with limited molecular characterization. Using cellular markers and knowledge on cellular sources of basement membrane laminins, we here describe molecularly distinct compartments surrounding different vessel types and provide a comprehensive characterization of the arachnoid and pial compartments and their connection to CNS vessels and perivascular pathways. We show that differential expression of plectin, E-cadherin and laminins α1, α2, and α5 distinguishes pial and arachnoid layers at the brain surface, while endothelial and smooth muscle laminins α4 and α5 and smooth muscle actin differentiate between arterioles and venules. Tracer studies reveal that interconnected perivascular compartments exist from arterioles through to veins, potentially providing a route for fluid flow as well as the transport of large and small molecules.
Keywords: Basement membranes; cerebrospinal fluid; fluid flow; laminin; perivascular pathways.
Figures
References
-
- Hutchings M, Weller RO. Anatomical relationships of the pia mater to cerebral blood vessels in man. J Neurosurg 1986; 65: 316–325. - PubMed
-
- Ichimura T, Fraser PA, Cserr HF. Distribution of extracellular tracers in perivascular spaces of the rat brain. Brain Res 1991; 545: 103–113. - PubMed
-
- Krisch BL, H.; Oksche, A. The meningea compartments of ME and cortex. Comparative analysis in rat. Cell Tissue Res 1983; 228: 597–640. - PubMed
-
- Krisch BL, Leonhardt H, Oksche A. Compartments and perivascular arrangement of the meninges covering the cerebral cortex of the rat. Cell Tissue Res 1984; 238: 459–474. - PubMed
-
- Rennels ML, Gregory TF, Blaumanis OR, et al. Evidence for a ‘paravascular' fluid circulation in the mammalian central nervous system, provided by the rapid distribution of tracer protein throughout the brain from the subarachnoid space. Brain Res 1985; 326: 47–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
