

Role of enhanced receptor engagement in the evolution of a pandemic acute hemorrhagic conjunctivitis virus
- PMID: 29284752
- PMCID: PMC5777043
- DOI: 10.1073/pnas.1713284115
Role of enhanced receptor engagement in the evolution of a pandemic acute hemorrhagic conjunctivitis virus
Abstract
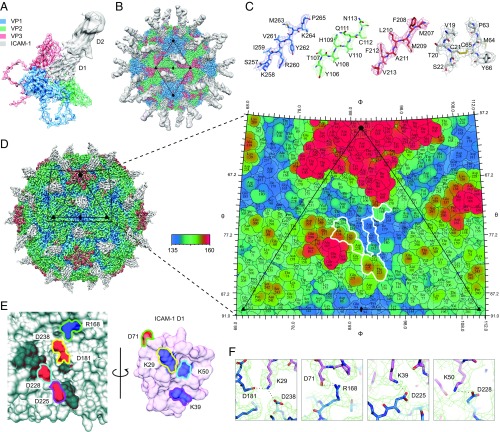
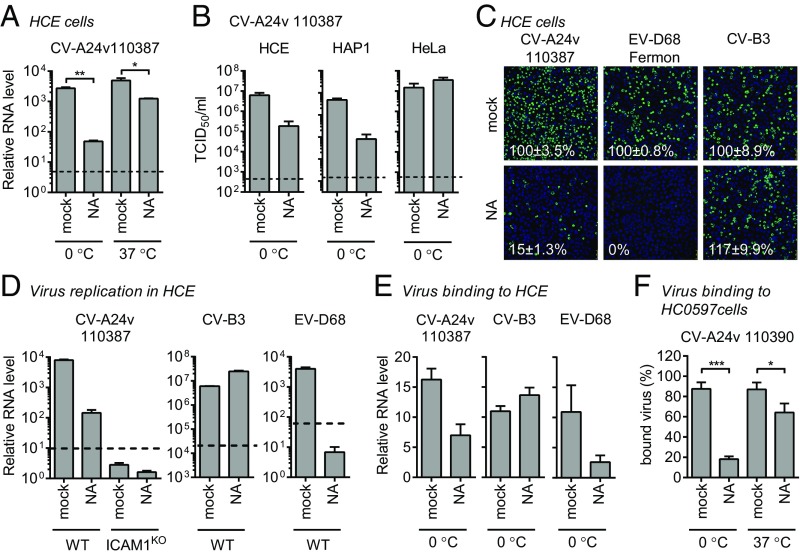
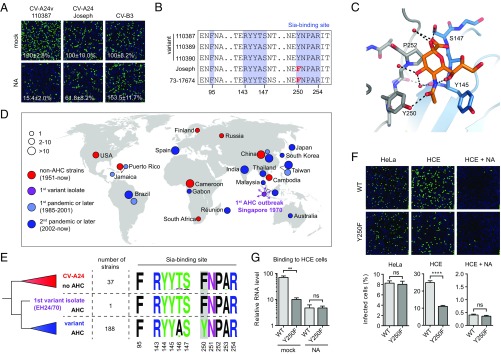
Acute hemorrhagic conjunctivitis (AHC) is a painful, contagious eye disease, with millions of cases in the last decades. Coxsackievirus A24 (CV-A24) was not originally associated with human disease, but in 1970 a pathogenic "variant" (CV-A24v) emerged, which is now the main cause of AHC. Initially, this variant circulated only in Southeast Asia, but it later spread worldwide, accounting for numerous AHC outbreaks and two pandemics. While both CV-A24 variant and nonvariant strains still circulate in humans, only variant strains cause AHC for reasons that are yet unknown. Since receptors are important determinants of viral tropism, we set out to map the CV-A24 receptor repertoire and establish whether changes in receptor preference have led to the increased pathogenicity and rapid spread of CV-A24v. Here, we identify ICAM-1 as an essential receptor for both AHC-causing and non-AHC strains. We provide a high-resolution cryo-EM structure of a virus-ICAM-1 complex, which revealed critical ICAM-1-binding residues. These data could help identify a possible conserved mode of receptor engagement among ICAM-1-binding enteroviruses and rhinoviruses. Moreover, we identify a single capsid substitution that has been adopted by all pandemic CV-A24v strains and we reveal that this adaptation enhances the capacity of CV-A24v to bind sialic acid. Our data elucidate the CV-A24v receptor repertoire and point to a role of enhanced receptor engagement in the adaptation to the eye, possibly enabling pandemic spread.
Keywords: ICAM-1; conjunctivitis; coxsackievirus A24v; receptor; sialic acid.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
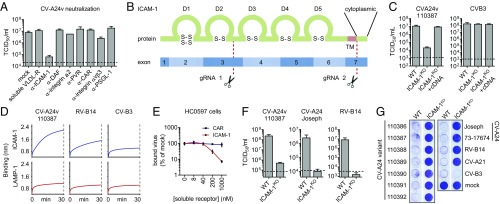
References
-
- Ghazali O, et al. An outbreak of acute haemorrhagic conjunctivitis in Melaka, Malaysia. Singapore Med J. 2003;44:511–516. - PubMed
-
- Kurokawa M, Rai SK, Ono K, Gurung R, Ruit S. Viral investigation of acute hemorrhagic conjunctivitis outbreak (2003) in Nepal using molecular methods. Southeast Asian J Trop Med Public Health. 2006;37:904–910. - PubMed
-
- Measroch V, Gear J, Faerber GI. Studies in poliomyelitis; the isolation of a coxsackie-like virus from the faeces of apparently healthy bantu infants. S Afr Med J. 1951;25:421–424. - PubMed
-
- Kono R, Sasagawa A, Ishii K, Sugiura S, Ochi M. Pandemic of new type of conjunctivitis. Lancet. 1972;1:1191–1194. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
