

Microglial Ramification, Surveillance, and Interleukin-1β Release Are Regulated by the Two-Pore Domain K+ Channel THIK-1
- PMID: 29290552
- PMCID: PMC5783715
- DOI: 10.1016/j.neuron.2017.12.002
Microglial Ramification, Surveillance, and Interleukin-1β Release Are Regulated by the Two-Pore Domain K+ Channel THIK-1
Abstract
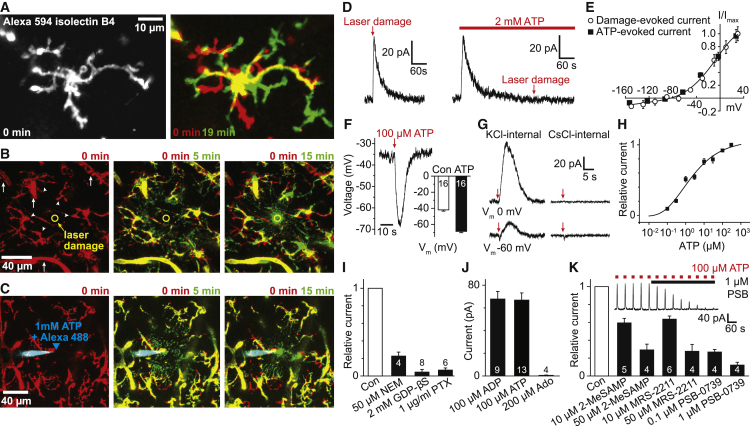
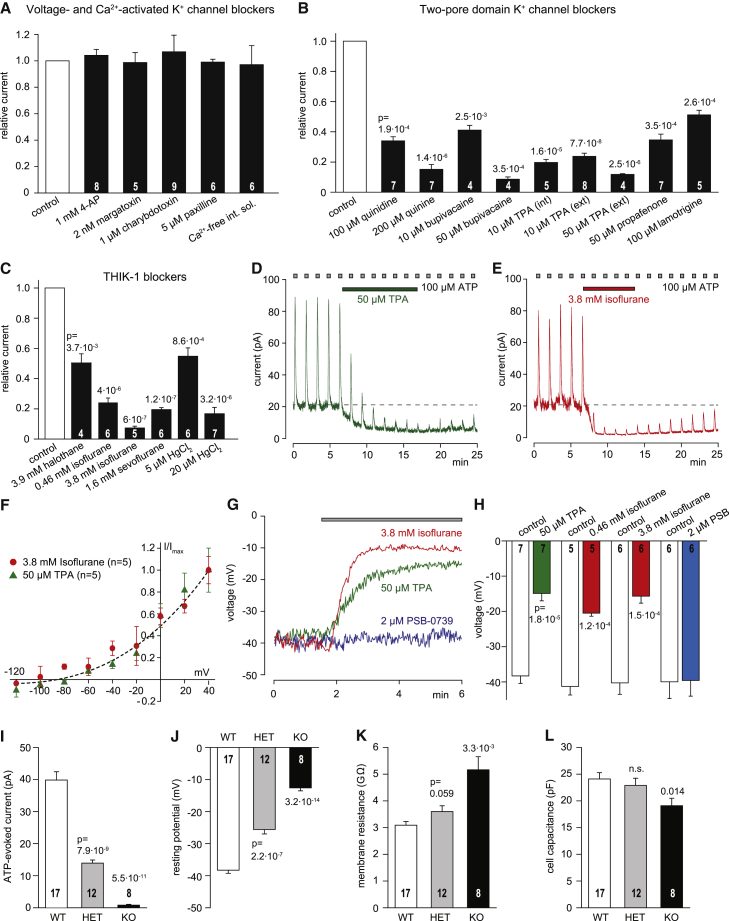
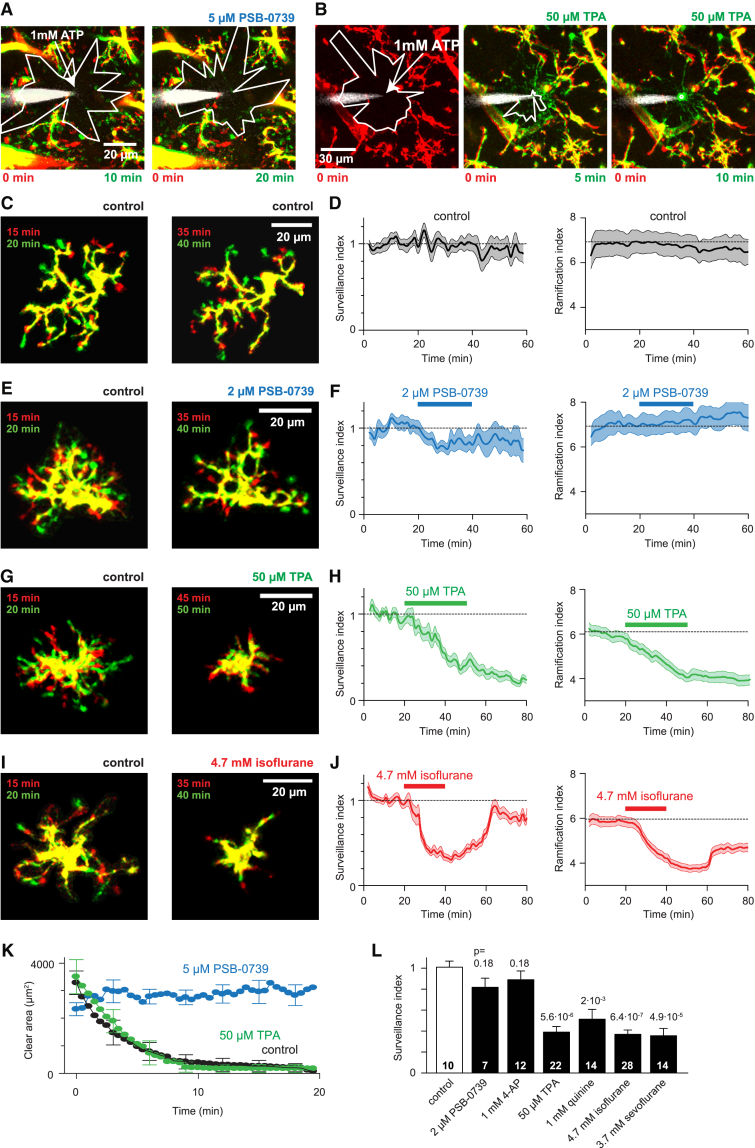
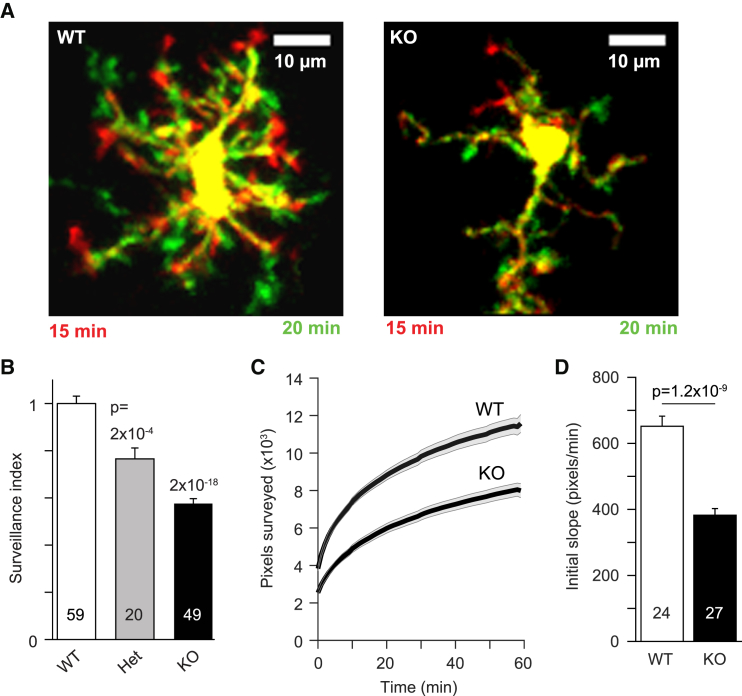
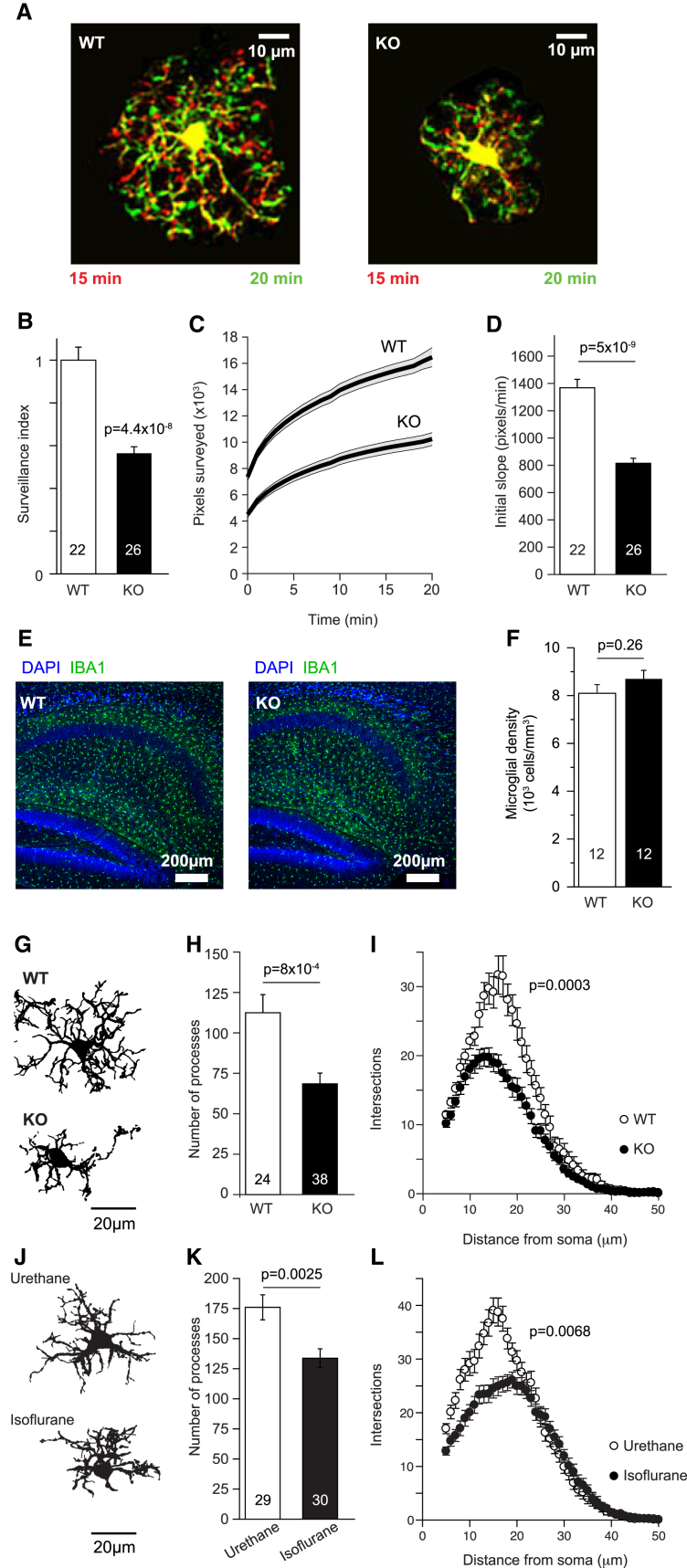
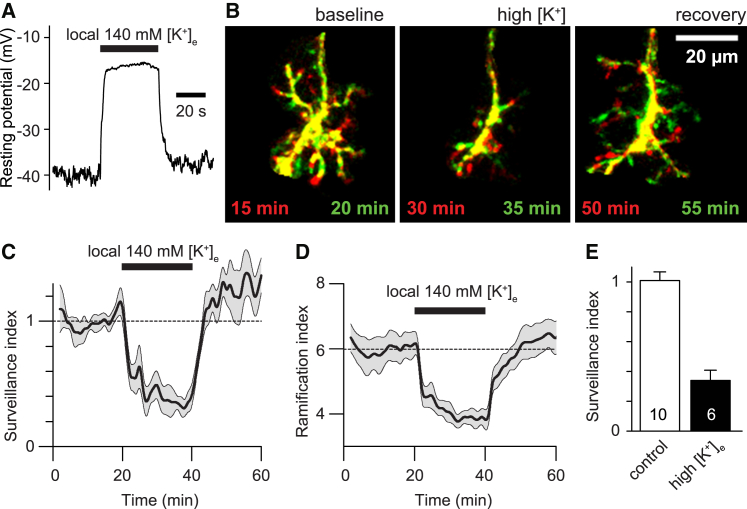
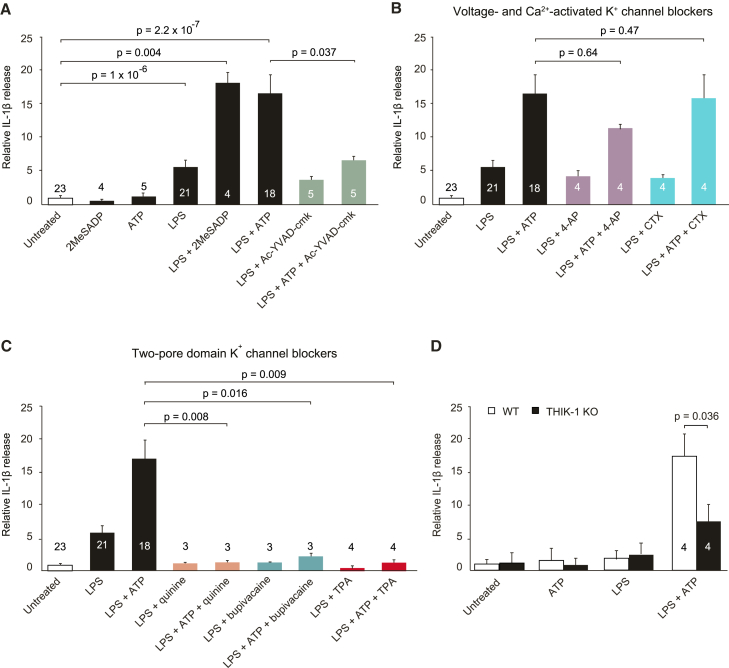
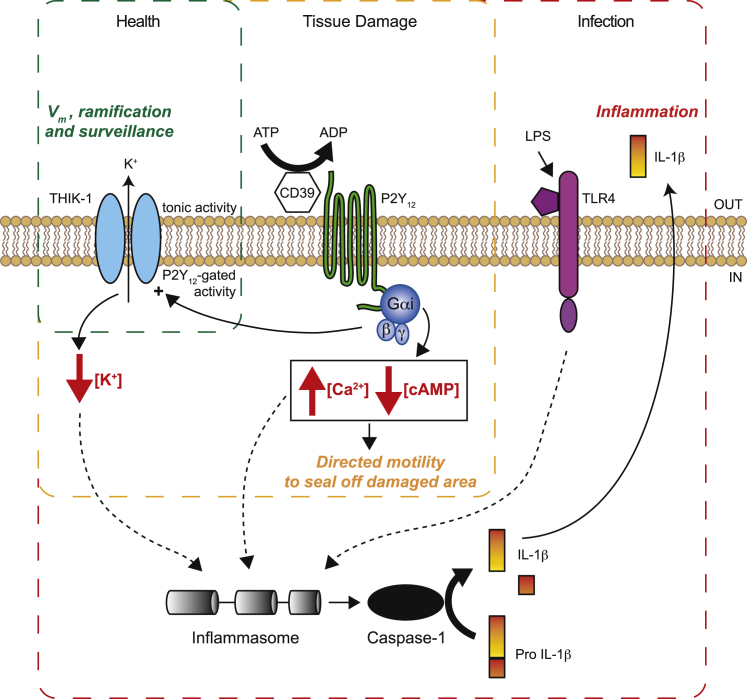
Microglia exhibit two modes of motility: they constantly extend and retract their processes to survey the brain, but they also send out targeted processes to envelop sites of tissue damage. We now show that these motility modes differ mechanistically. We identify the two-pore domain channel THIK-1 as the main K+ channel expressed in microglia in situ. THIK-1 is tonically active, and its activity is potentiated by P2Y12 receptors. Inhibiting THIK-1 function pharmacologically or by gene knockout depolarizes microglia, which decreases microglial ramification and thus reduces surveillance, whereas blocking P2Y12 receptors does not affect membrane potential, ramification, or surveillance. In contrast, process outgrowth to damaged tissue requires P2Y12 receptor activation but is unaffected by blocking THIK-1. Block of THIK-1 function also inhibits release of the pro-inflammatory cytokine interleukin-1β from activated microglia, consistent with K+ loss being needed for inflammasome assembly. Thus, microglial immune surveillance and cytokine release require THIK-1 channel activity.
Keywords: ATP; THIK-1; inflammasome; interleukin-1β; microglia; potassium channel; ramification; surveillance.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
The THIK and Thin of Microglia Dynamics.Neuron. 2018 Jan 17;97(2):253-255. doi: 10.1016/j.neuron.2018.01.002. Neuron. 2018. PMID: 29346745
-
Neuroimmunology: Brain police.Nat Rev Neurosci. 2018 Jan 19;19(2):60. doi: 10.1038/nrn.2018.5. Nat Rev Neurosci. 2018. PMID: 29348667 No abstract available.
References
-
- Bernardino L., Balosso S., Ravizza T., Marchi N., Ku G., Randle J.C., Malva J.O., Vezzani A. Inflammatory events in hippocampal slice cultures prime neuronal susceptibility to excitotoxic injury: a crucial role of P2X7 receptor-mediated IL-1beta release. J. Neurochem. 2008;106:271–280. - PubMed
-
- Bialas A.R., Presumey J., Das A., van der Poel C.E., Lapchak P.H., Mesin L., Victora G., Tsokos G.C., Mawrin C., Herbst R., Carroll M.C. Microglia-dependent synapse loss in type I interferon-mediated lupus. Nature. 2017;546:539–543. - PubMed
-
- Bischofberger J., Engel D., Li L., Geiger J.R.P., Jonas P. Patch-clamp recording from mossy fiber terminals in hippocampal slices. Nat. Protoc. 2006;1:2075–2081. - PubMed
-
- Boucsein C., Zacharias R., Färber K., Pavlovic S., Hanisch U.K., Kettenmann H. Purinergic receptors on microglial cells: functional expression in acute brain slices and modulation of microglial activation in vitro. Eur. J. Neurosci. 2003;17:2267–2276. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
