

Bacteria Facilitate Enteric Virus Co-infection of Mammalian Cells and Promote Genetic Recombination
- PMID: 29290575
- PMCID: PMC5764776
- DOI: 10.1016/j.chom.2017.11.007
Bacteria Facilitate Enteric Virus Co-infection of Mammalian Cells and Promote Genetic Recombination
Abstract
RNA viruses exist in genetically diverse populations due to high levels of mutations, many of which reduce viral fitness. Interestingly, intestinal bacteria can promote infection of several mammalian enteric RNA viruses, but the mechanisms and consequences are unclear. We screened a panel of 41 bacterial strains as a platform to determine how different bacteria impact infection of poliovirus, a model enteric virus. Most bacterial strains, including those extracted from cecal contents of mice, bound poliovirus, with each bacterium binding multiple virions. Certain bacterial strains increased viral co-infection of mammalian cells even at a low virus-to-host cell ratio. Bacteria-mediated viral co-infection correlated with bacterial adherence to cells. Importantly, bacterial strains that induced viral co-infection facilitated genetic recombination between two different viruses, thereby removing deleterious mutations and restoring viral fitness. Thus, bacteria-virus interactions may increase viral fitness through viral recombination at initial sites of infection, potentially limiting abortive infections.
Keywords: bacteria; co-infection; enteric virus; evolution; fitness; microbiota; poliovirus; recombination.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
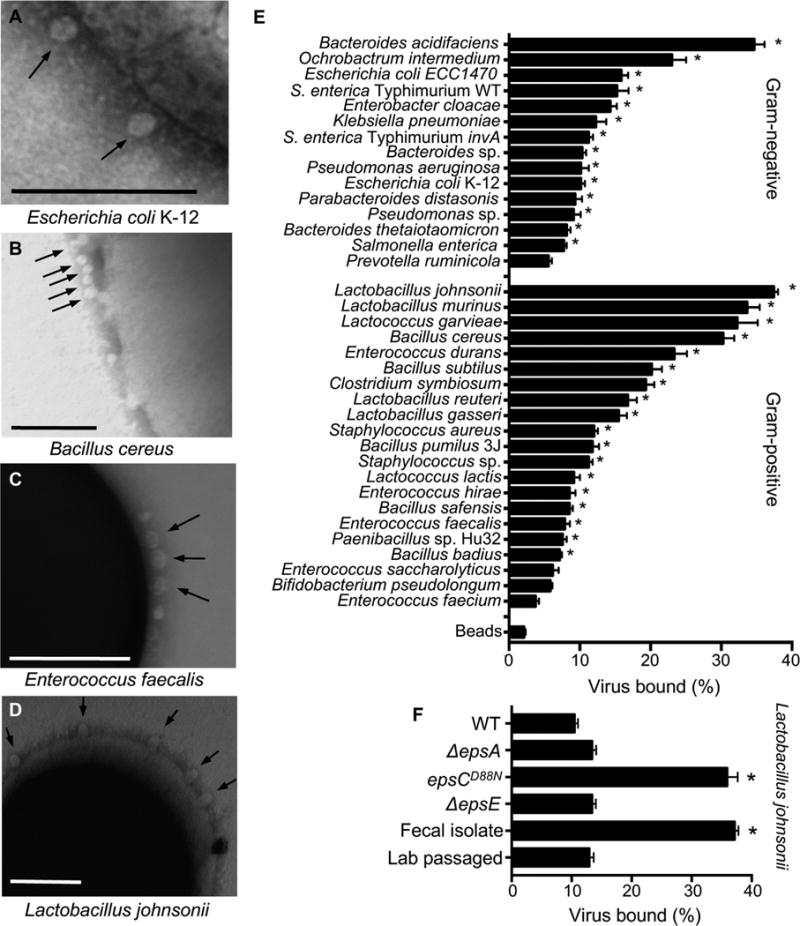
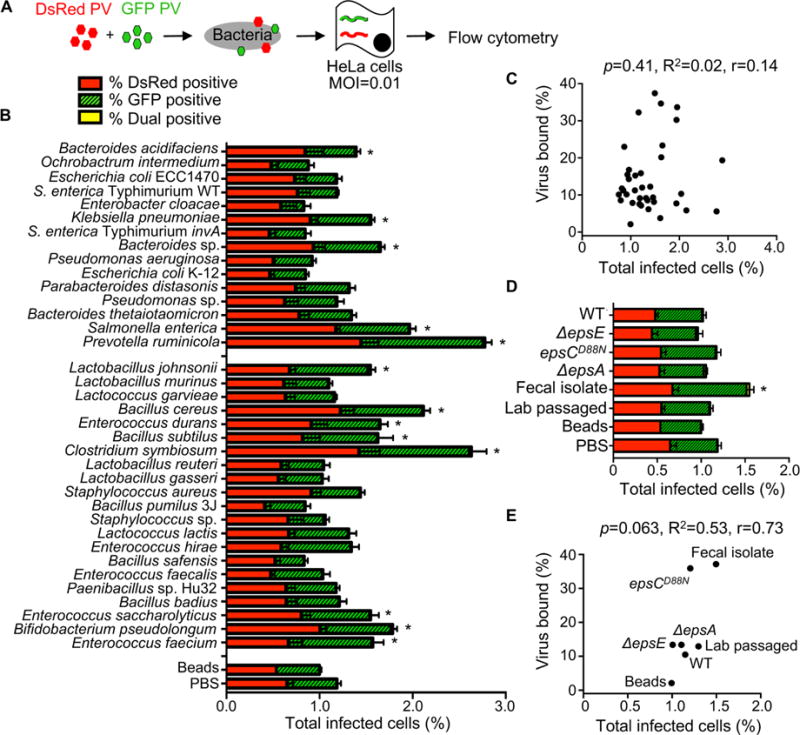
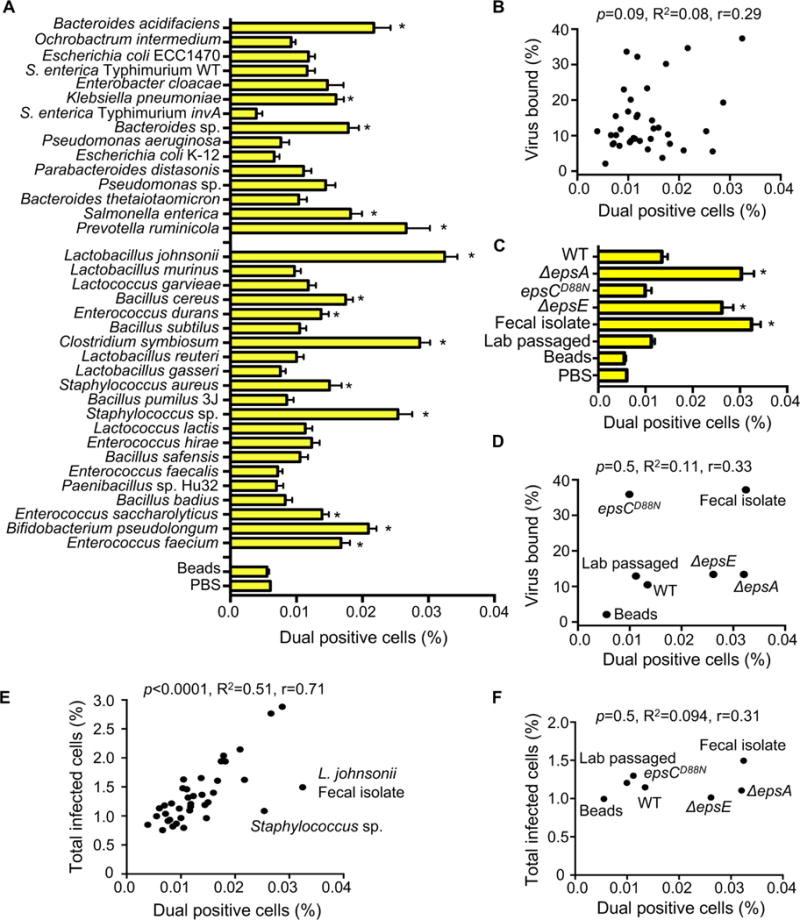
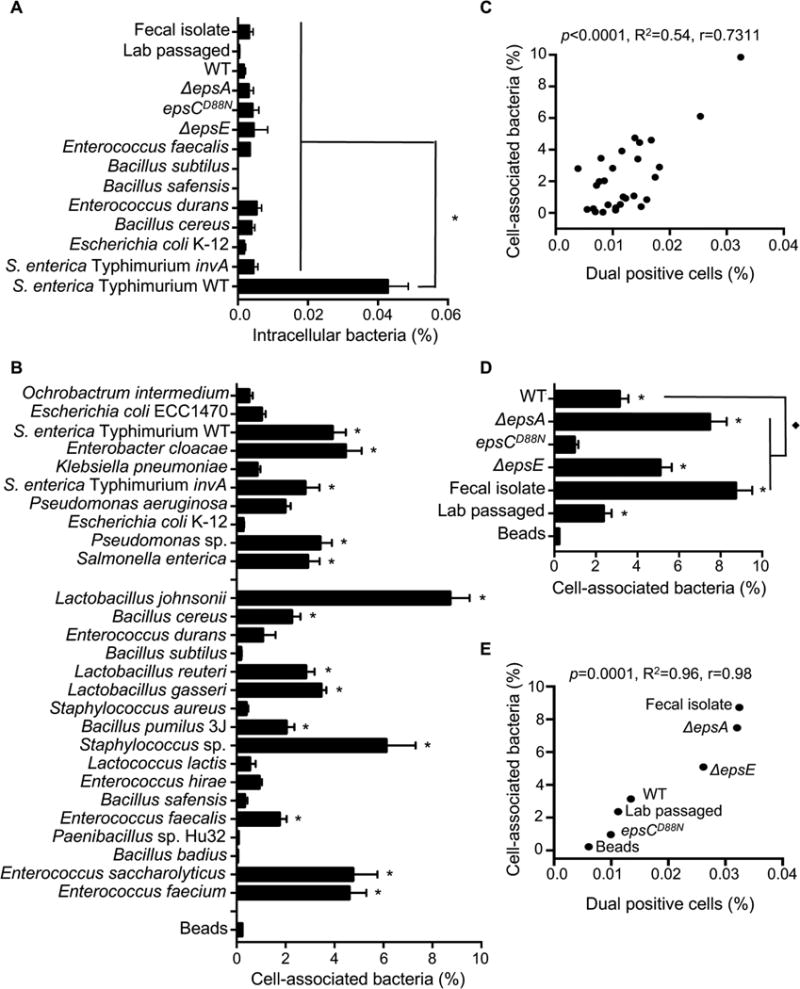

Comment in
-
Enteric Viruses Hitch a Ride on the Evolutionary Highway.Cell Host Microbe. 2018 Jan 10;23(1):5-6. doi: 10.1016/j.chom.2017.12.013. Cell Host Microbe. 2018. PMID: 29324229
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
